Определение частот аллелей и генотипов в популяциях по закону Харди-Вайнберга
Задача 76.
В стаде джейсейской породы крупного рогатого скота обследовали животных на наличие типов гемоглобина. Из 1200 обследованных животных с гемоглобином типа АА (HbAA) было 470, с гетерозиготным генотипом АВ (HbAB) – 700, с гомозиготным генотипом ВВ(HbBB) – 30 голов.
Определите частоту аллелей, ответственных за гемоглобин типа А и В и частоты генотипов в данном стаде.
Решение:
p – аллель гена А;
q – аллель гена В;
q2 генотип ВВ;
р2 – генотип АА;
2рq – генотип АВ.
1. Определим частоты генотипов в данном стаде, как в долях, так и в процентах, получим: генотип АА – (470/1200 = 0,392 или 39,2%), генотип Аа – (700/1200 = 0,583 или 58,3%), генотип ВВ – 0,025 (30/1200 = 0,025 или 2,5%).
2. Опираясь на формулу Харди-Вайнберга (р2 + 2рq + q2 = 1), вычислим частоту встречаемости аллеля (В) как корень квадратный из частоты встречаемости рецессивных гомозигот, получим: (q2 = 0,025; q = 0,158).
3. Учитывая, что суммарная частота встречаемости рецессивного и доминантного аллелей составляет 1 (р + q = 1), вычисляем частоту встречаемости аллеля доминантного гена (А), получим: 1 – 0,158 = 0,842.
4. Частоты генотипов в данном стаде определим, используя формулу Харди-Вайнберга, получим:
р2 + 2рq + q2 = 1;
(0,842)2 + (2 . 0,842 . 0,158) + (0,158)2 = 1;
0,709 + 0,266 + 0,025 = 1.
Ответ: частота аллеля (В) в популяции равна 0,158, частота аллеля (А) в популяции равна 0,842, частота генотипов (АА) равна 0,709, частота генотипов (АВ) равна 0,266, частота генотипов (ВВ) равна 0,025. Вновь полученные числа, рассчитанные по формуле, не совпадают с вычисленными изначально, кроме генотипа (аа), т.е. стадо не находится в генетическом равновесии и, очевидно, рецессивные гомозиготы не все выживают.
Задача 77.
Исходно в популяции частоты аллелей (А) и (а) равны 0,5. Как изменится частоты аллелей и генотипов в популяции в первом и втором поколениях, если:
а) Отбор действует против доминантных гомозигот, коэффициент отбора равен 0,5;
б) отбор действует против рецессивного фенотипа, коэффициент отбора равен 1?
Решение:
Для расчета частот генотипов под действием отбора преобразуют формулу Харди-Вайнберга с учетом этих величин.
Частоту аллеля (А) в F1 после отбора можно рассчитать по формуле:
p1 = [(p0)2 + p0q0]/[1 – S(q0)2], где
p0 – частота аллеля (А) в исходном поколении; q0 – частота аллеля (а) в исходном поколении, S – коэффициент отбора.
а) Отбор действует против доминантных гомозигот, коэффициент отбора равен 0,5.
1. Коэффициент отбора против доминантных гомозигот равный 0,5 говорит о том, что только половина их оставляют потомство. Исходя из этого, рассчитываем частоту аллеля (а) в первом поколении по формуле:
q1 = [(q0)2 + p0q0]/[1 – S(р0)2], где
p0 – частота аллеля (А) в исходном поколении; q0 – частота аллеля (а) в исходном поколении, S – коэффициент отбора.
Тогда
q1 = (0,25 + 0,25)/[1 – (0,5 . 0,25)] = 0,57
2. Рассчитываем частоту аллеля (A) в первом поколении, используя формулу:
р + q = 1;
p1 = 1- q1 = 1 – 0,57 = 0,43
3. По формуле Харди-Вайнберга рассчитываем частоты генотипов в популяции в первом поколении:
р2 + 2рq + q2 = 1;
(0,43)2 + (2 . 0,43 . 0,57) + (0,57)2 = 1;
0,19 + 0,49 + 0,32 = 1;
0,19(AA) + 0,49(Aa) + 0,32(aa) = 1;
(p1)2 = 0,19, 2р1q1 = 0,49, (q1)2 = 0,32.
4. Рассчитываем частоты аллелей и генотипов во втором поколении, получим:
q2 = [0,57 + (0,57 . 0,43)]/[1 – (0,5 . 0,32)] = 0,98;
p2 = 1 – q2 = 1 – 0,98 = 0,02;
(0,98)2 + (2 . 0,98 . 0,02) + (0,02)2 = 1;
0,9600 + 0,0396 + 0,0004 = 1;
0,9600(AA) + 0,0396(Aa) + 0,0004(aa) = ;
(p2)2 = 0,9600, 2р2q2 = 0,0396, (q2)2 = 0,0004.
Ответ: p1 = 0,43; q1 = 0,57; (p1)2 = 0,19; (p1)2 = 0,19; 2р1q1 = 0,49; (q1)2 = 0,32; p2 = 0,02; q2 = 0,98; (p2)2 = 0,9600; 2р2q2 = 0,0396; (q2)2 = 0,0004.
б) отбор действует против рецессивного фенотипа, коэффициент отбора равен 1.
1. Коэффициент отбора против рецессивных гомозигот равный 1 говорит о том, что все они не оставляют потомства. Исходя из этого, рассчитываем частоту аллеля A в первом поколении по формуле, получим:
p1 = [(p0)2 + p0q0]/[1 – S(q0)2], где
p0 – частота аллеля (А) в исходном поколении; q0 – частота аллеля (а) в исходном поколении, S – коэффициент отбора.
Тогда
p1 = (0,25 + 0,25) / (1 – 0,25) = 0,67
2. Рассчитываем частоту аллеля (а) в первом поколении:
q1 = 1- p1 = 1 – 0,67 = 0,33
3. По формуле Харди-Вайнберга рассчитываем частоты генотипов в популяции в первом поколении:
(0,67)2 + (2 . 0,67 . 0,33) + (0,33)2 = 1;
0,45(AA) + 0,44(Aa) + 0,11(aa) = 1.
4. Рассчитываем частоты аллелей и генотипов во втором поколении, получим:
p2 = (0,45 + 0,22)/(1 – 0,1) = 0,74;
q2 = 1- 0,74 = 0,26;
0,742 + (2 . 0,74 . 0,26) + 0,262 = 0,559(AA) + 0,39(Aa) + 0,06(aa) = 1.
Ответ: p1 = 0,67, q1 = 0,33, (p1)2 = 0, 45, 2p1q1 = 0,44, (q1)2 = 0,01; p2 = 0,74, q2 = 0,26, (p2)2 = 0, 55, 2p2q2 = 0,39, (q2)2 = 0,06.
Частоты аллелей
Любой
физический признак, например окраска
шерсти у мышей, определяется одним или
несколькими генами. Каждый ген может
существовать в нескольких различных
формах, которые называются аллелями
Число организмов в данной популяции,
несущих определенный аллель, определяет
частоту данного аллеля. Например, у
человека частота доминантного аллеля,
определяющего нормальную пигментацию
кожи, волос и глаз, равна 99%. Рецессивный
аллель, детерминирующий отсутствие
пигментации – так называемый альбинизм,
– встречается с частотой 1%. В популяционной
генетике частоту аллелей или генов
часто выражают не в процентах или в
простых дробях, а в десятичных дробях.
Таким образом, в данном случае частота
доминантного аллеля равна 0.99, а частота
рецессивного аллеля альбинизма – 0.01.
Общая частота аллелей в популяции
составляет 100%, или 1.0, поэтому
|
Частота доминантного аллеля |
+ |
Частота рецессивного аллеля |
=1 |
|
0.99 |
+ |
0.01 |
=1 |
Популяционная
генетика заимствовала у математической
теории вероятности два символа, p и q,
для выражения частоты, с которой два
аллеля, доминантный и рецессивный,
встречаются в генофонде данной популяции.
Таким образом,
p
+ q = 1,
где
p – частота доминантного, а q – частота
рецессивного аллеля.
В
примере с пигментацией у человека p =
0.99, а q = 0.01;
p
+ q = 1
0.99
+ 0.01 = 1
Значение
этого уравнения состоит в том, что, зная
частоту одного из аллелей, можно
определить частоту другого. Пусть,
например, частота рецессивного аллеля
= 25%, или 0.25. Тогда
p
+ q = 1
p
+ 0.25 = 1
p
= 1 – 0.25
p
= 0.75
Таким
образом, частота доминантного аллеля
равна 0.75, или 75%
1.3 Частота генотипов
Частоты
отдельных аллелей в генофонде позволяют
вычислять генетические изменения в
данной популяции и определять частоту
генотипов. Поскольку генотип данного
организма – главный фактор, определяющий
его фенотип, вычисление частоты генотипа
используют для предсказания возможных
результатов тех или иных скрещиваний.
Это имеет важное практическое значение
в сельском хозяйстве и медицине.
Математическая
зависимость между частотами аллелей и
генотипов известную под названием
равновесия Харди-Вайнберга, можно
сформулировать так: частоты доминантного
и рецессивного аллелей в данной популяции
будут оставаться постоянными из поколения
в поколение при наличии определенных
условий. Условия эти следующие:
1)
размеры популяции велики;
2)
спаривание происходит случайным образом;
3)
новых мутаций не возникает;
4)
все генотипы одинаково плодовиты, т.е.
отбора не происходит;
5)
поколения не перекрываются;
6)
не происходит ни эмиграции, ни иммиграции,
т.е. отсутствует обмен генами с другими
популяциями.
Поэтому
любые изменения частоты аллелей должны
быть обусловлены нарушением одного или
нескольких из перечисленных выше
условий. Все эти нарушения способны
вызвать эволюционное изменение; и если
такие изменения происходят, то изучать
их и измерять их скорость можно с помощью
уравнения Харди-Вайнберга.
Закон
Харди —
Вайнберга —
это закон популяционной
генетики —
в популяции
бесконечно большого размера, в которой
не действует отбор,
не идет мутационный
процесс, отсутствует обмен особями с
другими популяциями, не происходит
дрейф
генов,
все скрещивания случайны — частоты
генотипов
по какому-либо гену (в случае если в
популяции есть два аллеля этого гена)
будут поддерживаться постоянными из
поколения в поколение и соответствовать
уравнению:
p²
+ 2pq + q² = 1
Где
p² —
доля гомозигот по одному из аллелей;
p —
частота этого аллеля; q² —
доля гомозигот по альтернативному
аллелю; q —
частота соответствующего аллеля; 2pq —
доля гетерозигот.
Билет
№56 Элементарные эволюционные факторы:
мутации, популяционные волны,
генетико-автоматические процессы (дрейф
генов); их значение в изменении
генотипической структуры популяций
Мутации
– это внезапные скачкообразные стойкие
изменения в структуре генотипа. Организмы,
у которых произошла мутация, называются
мутантами. При существенном изменении
условий существования те мутации,
которые раньше были вредными, могут
оказаться полезными. Таким образом,
мутации являются материалом для
естественного
отбора.
Популяционные
волны
– это вспышки численности, периодические
или непериодические значительные
изменения числа особей в популяции.
Популяционные волны вместе с мутационными
процессами являются фактором-поставщиком
эволюционного материала, выводя ряд
генотипов, совершенно случайно и
ненаправленно, в качестве «кандидатов»
на роль новых звеньев в протекающих
эволюционных явлениях и процессах.
Давление этого фактора может быть весьма
различно и, вероятно, обычно превышает
таковое мутационного процесса.
ДРЕЙФ
ГЕНОВ
(генетико-автоматические процессы) —
один из факторов генетической динамики
популяции, чье действие приводит к
изменению ее генотипической структуры.
Дрейф генов — это изменение частоты
гена в популяции под действием случайных
причин. К их числу относят колебания
численности популяции, ее возрастного
и полового состава, изменение кормовой
базы, наличие или отсутствие конкуренции
и т.п. Основной генетический эффект
дрейфа — постепенная гомозиготизация
популяции. Происходит некоторое
скрещивание с соседними популяциями,
что может приводить к значительному
обмену генами; в популяцию проникают
новые гены, генетическая изменчивость
ее возрастает, и это может играть
известную роль в эволюции данного вида
или отдельной популяции.
Билет№57
Генетический полиморфизм и наследственное
разнообразие природных популяций. Формы
полиморфизма (адаптационный и
балансированный). Генетический груз и
его эволюционное значение.
Полиморфизм.
Популяция
называется полиморфной по данному
локусу, если в ней встречается два или
большее число аллелей. Исследуя много
локусов, можно определить среди них
долю полиморфных, т.е. оценить степень
полиморфизма,
которая является показателем генетического
разнообразия популяции. Под биологическим
разнообразием понимается все «множество
различных живых организмов, изменчивость
среди них и экологических комплексов,
частью которых они являются, что включает
разнообразие внутри видов, между видами
и экосистемами»; при этом следует
различать глобальное и локальное
разнообразие. Биологическое разнообразие
представляет собой один из важнейших
биологических ресурсов (биологическим
ресурсом считается «генетический
материал, организмы или их части, либо
экосистемы, используемые или потенциально
полезные для человечества, включая
природное равновесие внутри экосистем
и между ними»). Наличие двух и более
аллелей (а, соответственно, и генотипов)
в популяции называется генетическим
полиморфизмом.
Условно принято, что частота наиболее
редкого аллеля при полиморфизме должна
быть не менее 1% (0,01). Существование
генетического полиморфизма – обязательное
условие сохранения биоразнообразия.
Адаптационный
полиморфизм
возникает, если в различных, но закономерно
изменяющихся условиях жизни отбор
благоприятствует разным генотипам.
Так, в популяциях двухточечных божьих
коровок Adalia
bipunctata при
уходе на зимовку преобладают черные
жуки, а весной—красные (рис. 11.7). Это
происходит потому, что красные формы
лучше переносят холод, а черные интенсивнее
размножаются в летний период.
Балансированный
полиморфизм
возникает, если отбор благоприятствует
гетерозиготам в сравнении с рецессивными
и доминантными гомозиготами. Так, в
опытной численно равновесной популяции
плодовых мух Drosophila
melanogaster,
содержащей поначалу много мутантов с
более темными телами (рецессивная
мутация ebony), концентрация последних
быстро падала, пока не стабилизировалась
на уровне 10% (рис. 11.8). Анализ показал,
что в созданных условиях гомозиготы по
мутации ebony и гомозиготы по аллелю дикого
типа менее жизнеспособны, чем гетерозиготные
мухи. Это и создает состояние устойчивого
полиморфизма по соответствующему
локусу.
Существование
в популяции неблагоприятных аллелей в
составе гетерозиготных генотипов
называют генетическим
грузом. Как
отмечалось в разделе 1.5, некоторые
рецессивные аллели, вредоносные в
гомозиготном состоянии могут сохраняться
в гетерозиготных генотипах и при
некоторых условиях среды доставлять
селективное преимущество; примером
служит аллель серповидноклеточности
в местах распространения малярии. Любое
повышение частоты рецессивных аллелей
в популяции в результате вредных мутаций
увеличивает ее генетический груз.
Билет
№58 Популяционная структура человечества.
Демографическая характеристика. Роль
системы браков в распределении аллелей
в популяции. Использование закона
Харди-Вайнберга в характеристике
генетической структуры популяции людей.
Популяция
– это группа людей, занимающих определённую
территорию и свободно вступающих в
брак. Обычно под популяцией понимают
как население города, района, так и
определённой местности, используя этот
термин в экологическом или географическом
контекстах.Внутри популяции браки
заключаются чаще, чем с членами других
популяций. Это обеспечивает поддержание
генофонда. При сохранении в его пределах
генетической гетерогенности, резерва
наследственной изменчивости
обеспечивается
приспособляемость популяции к факторам
среды. С развитием человечества, с
возникновением социальных отношений
в обществе, популяция
человека
включается в систему специфических для
людей социальных структур. Популяция
человека в этих условиях зависит от
природных и социальных факторов и
оказывает на них влияние.
На
начальных этапах человеческой истории
на основе сложных взаимоотношений между
смежными популяциями начали образовываться
человеческие расы.
В экологическом
смысле раса – более или менее искусственное
понятие, основанное для многих популяций
на выделении их из различных экосистем
и
объединении на основе общности или
частоты появления некоторого числа
физиологических признаков. Люди разных
рас по всем основным биологическим и
психологическим особенностям близки
друг другу и принадлежат к одному виду
– человек разумный.По мере развития
экономического, социального и
биологического взаимодействия, усиления
миграционных процессов возникают
сообщества людей с
разнообразным
сочетанием расовых признаков, а нередко
наблюдается стирание расовых различий.
Роль
системы браков в распределении аллелей
в популяции
Для
характеристики популяции важным является
определение её как репродуктивного
сообщества людей, обладающих общим
генетическим фондом. Поэтому в формировании
популяции людей главную роль играет не
общность территории, а родственные
связи между особями популяции.
Демографическими
показателями
популяции людей служат размер, уровень
рождаемости и смертности, возрастная
структура, экономическое состояние,
уклад жизни и т.п.
В
популяции часто образуются более мелкие
группировки людей,изолированных с точки
зрения размножения (лишённых возможности
свободно заключать брачные союзы в
пределах популяции). Изолирующими
факторами могут выступать географические
(большое расстояние или препятствие к
передвижению), но чаще всего факторы
социального или религиозного порядка.
При этом жители даже одного небольшого
района могут часто образовывать ряд
совершенно изолированных групп людей
(изолятов) численностью до 1500 человек,
но чаще всего численностью в несколько
десятков особей. Закрепляющиеся гены
имеют тенденцию ограничиваться именно
этой группой (частота внутригрупповых
браков в изолятах превышает 90%). Члены
изолятов через 4 поколения (примерно
через 100 лет) являются уже по крайней
мере троюродными сибсами. Популяции
людей численностью от 1500 до 4000 человек
называют демами. Частота внутригрупповых
браков в демах составляет 80-90%. В ранние
периоды истории человечества расстояния
и другие географические особенности
служили наиболее устойчивыми барьерами
на пути к смешению демов и изолятов, в
чём кроется одна из причин широкой
географической вариабельности человека
как вида.
Строгой
панмиксии в популяциях людей не
существует: предпочтения в выборе
брачного партнёра и социальные факторы
препятствуют подлинно случайному
заключению браков. Тем не менее существуют
гены (например, определяющие группы
крови), которые обычно редко принимаются
во внимание при подборе брачных пар и
распределяются среди жителей популяции
так, как если бы подбор брачных пар
происходил более или менее случайно.Однако
в настоящее время в связи с развитием
транспорта круг возможных браков
значительно расширился, замкнутость
изолятов нарушается во всех частях
Земли и, по-видимому, навсегда.
Закон
Харди–Вайнберга – основной закон
популяционной генетики
Структура
генофонда в панмиктической стационарной
популяции описывается основным законом
популяционной генетики – законом
Харди-Вайнберга,
который гласит, что в
идеальной популяции существует постоянное
соотношение относительных частот
аллелей и генотипов, которое описывается
уравнением:
(p
A
+ q a)2
=
р2
АА
+ 2∙р∙q
Aa
+ q2
aa
= 1
Практическое
значение закона Харди–Вайнберга
1.
В
здравоохранении
– позволяет оценить популяционный
риск
генетически обусловленных заболеваний
2.
В
селекции
– позволяет выявить генетический
потенциал
исходного материала (природных популяций,
а также сортов и пород народной селекции
3.
В
экологии
– позволяет выявить влияние самых
разнообразных факторов на популяции.
Вопрос
№59 Особенности действия элементарных
эволюционных факторов в человеческих
популяциях (мутации, миграции). Мутационный
груз, его биологическая сущность и
значение. Опасность индуцированного
мутагенеза.
Человеческие
мутации,миграции
В
основе мутаций лежат наследуемые
изменения генетического материала. В
результате мутации возникает мутантная
аллель гена или мутантная хромосома,
обусловливающие появление мутантного
признака. Мутации могут возникнуть в
любой момент, но их появление более
вероятно в делящейся, а не в покоящейся
клетке. В генетическом отношении важны
те мутации, которые возникают при
гаметогенезе и наследуются особями
потомства. Частота мутации для единичного
локуса составляет в среднем 1:100000 половых
клеток, однако у человека в целом, мутация
вовсе не редкое явление.
Мутагенами
по отношению к человеку выступают не
только естественные факторы
(ультрафиолетовое излучение, температура,
ионизирующее излучение, определённая
химическая среда), но и факторы, производные
научно-технического прогресса
(рентгеновские излучения и другие
физические факторы, синтетические смолы
и другие химические вещества). На частоту
мутаций у человека оказывает влияние
возраст. Некоторые гены Х-хромосомы
мутируют в мужском организме чаще, чем
в женском.
Генотип
человека – это высокоинтегрированная
система взаимодействующих генов (а
также составляющих их элементов), и
случайные изменения в её составе влияют
на неё чаще всего отрицательно. Поэтому
большинство мутантных генов оказываются
вредными для человека.
В
небольших популяциях людей мутантные
гены могут сохраняться или утрачиваться
случайным образом. В них хорошо выражен
дрейф генов
– изменение частоты генов в популяции
в ряду поколений под действием чисто
случайных факторов. В настоящее время
общепризнанным является тот факт, что
различия в частоте некоторых групп
крови между близкими поселениями людей
в отдалённых уголках земного шара
возникли вследствие дрейфа генов
Обобщённым примером влияния дрейфа
генов на частоту аллелей является
«эффект родоначальника». Он возникает,
когда несколько семей выселяются на
новую территорию и поддерживают высокий
уровень брачной изоляции, порвав
практически все связи с родительской
популяцией. В этом случае в генофонде
переселенцев из-за небольшой численности
особей случайно закрепляются одни
аллели и элиминируются другие.
Последствиями дрейфа генов, очевидно,
является неравномерное распределение
некоторых наследственных заболеваний
по группам населения земного
шараСущественное влияние на генофонды
популяций людей оказывал фактор изоляции.
Длительным проживанием в состоянии
относительной географической и культурной
изоляции объясняют, например, некоторые
антропологические особенности
представителей малых народностей:
своеобразный рельеф ушной раковины
бушменов, большую ширину нижнечелюстного
диаметра коряков и ительменов,
исключительное развитие бороды у айнов.
Сохранению
высокого уровня генетической изоляции
двух популяций (изолятов), существующих
на одной территории, способствуют
отличия по физическим признакам или
образу жизни. Однако такие барьеры в
последнее время исчезают, о чём
свидетельствует, например, тот факт,
что доля генов от белых людей возросла
в настоящее время у американских негров
до 25%, а у бразильских негров – до 40%.,это
связано с процессом Миграции особей.
Мутационный
груз —
снижение средней приспособленности
популяции за счет непрерывного
возникновения мутаций, снижающих
жизнеспособность особей. В результате
чего происходит изменение в генотипе
каждой особи данной популяции, снижается
численность вида, а если мутация является
полезной в конкретных условиях, то
наоборот происходит резкое увеличение
популяции.
Индуцированный
мутагенез
вызывается различными мутагенами; для
человеческих популяций особенно опасен.
Экспериментальные разработки в области
индуцированного мутагенеза у человека
являются основой оценки вредных факторов
внешней среды с генетических позиций
и их гигиенического нормирования.
Мутагенез определяет возможность
спонтанных абортов, врождённых пороков
развития, наследственных болезней с
чёткими клиническими проявлениями,
хромосомных болезней и др.
Вопрос
№60 Специфика действия естественного
отбора и изоляции в генетических
популяциях. Демы. Изоляты. Дрейф генов.
Особенности генофондов изолятов.
Специфика
действия естественного отбора
Одним
из наиболее важных эволюционных факторов,
изменяющих частоты аллельных генов в
популяциях людей, является естественный
отбор. Однако его давление в человеческих
популяциях ослабело настолько, что
отбор утратил значение как фактор
видообразования. Это обусловлено
возрастанием значения социальных
факторов исторического развития
человечества и постепенным ослаблением
роли биологических факторов эволюции
человека.
Однако
за естественным отбором осталась функция
стабилизации генофондов и поддержания
наследственного разнообразия популяций
людей. О действии на популяцию человека
стабилизирующей формы естественного
отбора свидетельствует,большая
перинатальная смертность среди
недоношенных и переношенных новорождённых.
Направление отбора в этом случае
определяется снижением общей
жизнеспособности новорождённых.
Отрицательное
действие отбора по одному локусу
иллюстрирует наследование антигенов
системы резус. Такой направленный против
гетерозигот отбор приводит к уменьшению
частоты более редкого (рецессивного,
d) аллеля в европейской популяции.
Несовместимость матери и плода характерна
также для системы групп крови AB0, которая,
в отличие от резус-иммунизации, может
сказаться уже на первом ребёнке. Для
системы AB0 существуют и другие селективные
факторы: в настоящее время продемонстрирована
ассоциация групп крови AB0 с очень многими
заболеваниями например, лица с группой
крови А чаще заболевают раком, тогда
как лица, имеющие группу крови 0, более
подвержены язве желудка и двенадцатиперстной
кишки; а риск заболевания ревматизмом
самый низкий среди лиц групп 0. Эти
селективные факторы влияют на частоту
аллелей, определяющих группы крови.
Особой
жёсткостью выделяется отбор, направленный
против Действию отбора, снижающего в
генофондах некоторых популяций людей
концентрацию определённых аллелей,
может противостоять контротбор, который,
наоборот, поддерживает частоту этих
аллелей на достаточно высоком
уровнеГенофонд популяций человека
является результатом наложения
многочисленных и разнонаправленных
векторов отбора, обеспечивающего
сохранение в каждом поколении сравнительно
приспособленных к данным условиям
генотипов. При этом с течением времени
влияние отбора на генетическую структуру
популяций людей снижается в основном
благодаря успехам лечебной и
профилактической медицины, а также
социально-экономическим преобразованиям
цивилизации.
Демы.
Изоляты
В
популяции часто образуются более мелкие
группировки людей,изолированных с точки
зрения размножения (лишённых возможности
свободно заключать брачные союзы в
пределах популяции). Изолирующими
факторами могут выступать географические
(большое расстояние или препятствие к
передвижению), но чаще всего факторы
социального или религиозного порядка.
При этом жители даже одного небольшого
района могут часто образовывать ряд
совершенно изолированных групп людей
(изолятов) численностью до 1500 человек,
но чаще всего численностью в несколько
десятков особей. Закрепляющиеся гены
имеют тенденцию ограничиваться именно
этой группой (частота внутригрупповых
браков в изолятах превышает 90%). Члены
изолятов через 4 поколения (примерно
через 100 лет) являются уже по крайней
мере троюродными сибсами. Популяции
людей численностью от 1500 до 4000 человек
называют демами. Частота внутригрупповых
браков в демах составляет 80-90%. В ранние
периоды истории человечества расстояния
и другие географические особенности
служили наиболее устойчивыми барьерами
на пути к смешению демов и изолятов, в
чём кроется одна из причин широкой
географической вариабельности человека
как вида.
дрейф
генов
– изменение частоты генов в популяции
в ряду поколений под действием чисто
случайных факторов. В настоящее время
общепризнанным является тот факт, что
различия в частоте некоторых групп
крови между близкими поселениями людей
в отдалённых уголках земного шара
возникли вследствие дрейфа генов
Билет
№61 Генетический полиморфизм – основа
внутри- и межпопуляционной изменчивости
человека. Значение полиморфизма в
предрасположенности к заболеваниям, к
реакциям на аллергены, лекарственные
препараты, пищевые продукты. Значение
генетического разнообразия в будущем
человечества.
Закон Харди-Вайнберга
Популяционная генетика занимается генетической структурой популяций.
Понятие «популяция» относится к совокупности свободно скрещивающихся особей одного вида, длительно существующей на определенной территории (части ареала) и относительно обособленной от других совокупностей того же вида.
Важнейший признак популяции – это относительно свободное скрещивание. Если возникают какие-либо изоляционные барьеры, препятствующие свободному скрещиванию, то возникают новые популяции.
У человека, например, помимо территориальной изоляции, достаточно изолированные популяции могут возникать на основе социальных, этнических или религиозных барьеров. Поскольку между популяциями не происходит свободного обмена генами, то они могут существенно различаться по генетическим характеристикам. Для того чтобы описывать генетические свойства популяции, вводится понятие генофонда: совокупности генов, встречающихся в данной популяции. Помимо генофонда важны также частота встречаемости гена или частота встречаемости аллеля.
Знание того, как реализуются законы наследования на уровне популяций, принципиально важно для понимания причин индивидуальной изменчивости. Все закономерности, выявляемые в ходе психогенетических исследований, относятся к конкретным популяциям. В других популяциях, с иным генофондом и другими частотами генов, могут получаться отличающиеся результаты.
Закон Харди-Вайнберга— основа математических построений генетики популяций и современной эволюционной теории. Сформулирован независимо друг от друга математиком Г. Харди (Англия) и врачом В. Вайнбергом (Германия) в 1908 г. Этот закон утверждает, что частоты аллелей и генотипов в данной популяции будут оставаться постоянными из поколения в поколение при выполнении следующих условий:
1) численность особей популяции достаточно велика (в идеале — бесконечно велика),
2) спаривание происходит случайным образом (т. е. осуществляется панмиксия),
3) мутационный процесс отсутствует,
4) отсутствует обмен генами с другими популяциями,
5) естественный отбор отсутствует, т. е. особи с разными генотипами одинаково плодовиты и жизнеспособны.
Иногда этот закон формулируют иначе: в идеальной популяции частоты аллелей и генотипов постоянны. (Поскольку описанные выше условия выполнения данного закона и есть свойства идеальной популяции.)
Математическая модель закона отвечает формуле:
p2+2pq+q2=1
Она выводится на основе следующих рассуждений. В качестве примера возьмем простейший случай — распределение двух аллелей одного гена. Пусть два организма являются основателями новой популяции. Один из них является доминантной гомозиготой (АА), а другой — рецессивной гомозиготой (аа). Естественно, что все их потомство в F1 будет единообразным и будет иметь генотип (Аа). Далее особи F1 будут скрещиваться между собой. Обозначим частоту встречаемости доминантного аллеля (А) буквой p, а рецессивного аллеля (а) — буквой q. Поскольку ген представлен всего двумя аллелями, то сумма их частот равна единице, т. е. р + q = 1. Рассмотрим все яйцеклетки в данной популяции. Доля яйцеклеток, несущих доминантный аллель (А), будет соответствовать частоте этого аллеля в популяции и, следовательно, будет составлять р. Доля яйцеклеток, несущих рецессивный аллель (а), будет соответствовать его частоте и составлять q. Проведя аналогичные рассуждения для всех сперматозоидов популяции, придем к заключению о том, что доля сперматозоидов, несущих аллель (А), будет составлять р, а несущих рецессивный аллель (а) — q. Теперь составим решетку Пеннета, при этом при написании типов гамет будем учитывать не только геномы этих гамет, но и частоты несомых ими аллелей. На пересечении строк и столбцов решетки мы получим генотипы потомков с коэффициентами, соответствующими частотам встречаемости этих генотипов.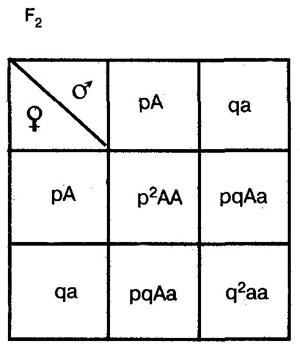
Из приведенной решетки видно, что в F2 частота доминантных гомозигот (АА) составляет р, частота гетерозигот (Аа) — 2pq, а рецессивных гомозигот (аа) — q. Поскольку приведенные генотипы представляют собой все возможные варианты генотипов для рассматриваемого нами случая, то сумма их частот должна равняться единице, т. е.
p2+2pq+q2=1
Главное применение закона Харди—Вайнберга в генетике природных популяций — вычисление частот аллелей и генотипов.
Рассмотрим пример использования этого закона в генетических расчетах. Известно, что один человек из 10 тыс. является альбиносом, при этом признак альбинизма у человека определяется одним рецессивным геном. Давайте вычислим, какова доля скрытых носителей этого признака в человеческой популяции. Если один человек из 10 тыс. является альбиносом, то это значит, что частота рецессивных гомозигот составляет 0,0001, т. е. q2 = 0,0001. Зная это, можно определить частоту аллеля альбинизма q, частоту доминантного аллеля нормальной пигментации р и частоту гетерозиготного генотипа (2pq). Люди с таким генотипом как раз и будут скрытыми носителями альбинизма, несмотря на то что фенотипически этот ген не будет у них проявляться и они будут иметь нормальную пигментацию кожи.
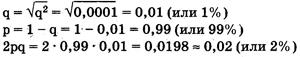
Из приведенных простых расчетов видно, что, хотя число альбиносов крайне невелико — всего лишь один человек на 10 тыс., ген альбинизма несет значительное количество людей — около 2% . Иными словами, даже если признак фенотипически проявляется очень редко, то в популяции присутствует значительное количество носителей этого признака, т. е. особей, имеющих этот ген в гетерозиготе.
Благодаря открытию закона Харди—Вайнберга процесс микроэволюции стал доступен непосредственному изучению: о его ходе можно судить по изменениям из поколения в поколение частот генов (или генотипов). Таким образом, несмотря на то что этот закон действителен для идеальной популяции, которой нет и не может быть в природе, он имеет огромное практическое значение, так как дает возможность рассчитать частоты генов, изменяющиеся под влиянием различных факторов микроэволюции.
ПРИМЕРЫ РЕШЕНИЯ ЗАДАЧ
1.Альбинизм у ржи наследуется как аутосомный рецессивный признак. На участке из 84000 растений 210 оказались альбиносами. Определить частоту гена альбинизма у ржи.
Решение
В связи с тем, что альбинизм у ржи наследуется как аутосомный рецессивный признак, все растения альбиносы будут гомозиготны по рецессивному гену — аа. Частота их в популяции (q2) равна 210/84000 = 0,0025. Частота рецессивного гена а будет равна 0,0025. Следовательно, q = 0,05.
Ответ:0,05
2.У крупного рогатого скота красная масть неполностью доминирует над белой (гибриды имеют чалую окраску). В районе обнаружены: 4169 красных, 756 белых и 3708 чалых животных. Какова частота генов окраски скота в этом районе?
Решение.
Если ген красной масти животных обозначить через А,
а ген белой — а, то у красных животных генотип будет АА (4169), у чалых Аа (3780), у белых – аа (756). Всего зарегистрировано животных 8705. Можно рассчитать частоту гомозиготных красных и белых животных в долях единицы. Частота белых животных будет 756 : 8705 = 0.09. Следовательно q2 =0.09 . Частота рецессивного гена q = 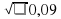 = 0,3. Частота гена А будет р = 1 — q. Следовательно, р = 1 – 0,3 = 0,7.
= 0,3. Частота гена А будет р = 1 — q. Следовательно, р = 1 – 0,3 = 0,7.
Ответ: р = 0,7, гена q = 0,3.
3.У человека альбинизм – аутосомный рецессивный признак. Заболевание встречается с частотой 1 / 20 000. Определите частоту гетерозиготных носителей заболевания в районе.
Решение.
Альбинизм наследуется рецессивно. Величина 1/20000 –
это q2. Следовательно, частота гена а будет: q = 1/20000 =
= 1/141. Частота гена р будет: р = 1 – q; р = 1 – 1/141 = 140/141.
Количество гетерозигот в популяции равно 2pq = 2 х (140/141) х (1/141) = 1/70. Т.к. в популяции 20000 человек то число гетерозигот в ней 1/ 70 х 20000 = 286 человек.
Ответ: 286 человек
4.Врожденный вывих бедра у человека наследуется как сутосомный доминантный признак с пенетрантностью 25%. Болезнь встречается с частотой 6:10 000. Определите число гетерозиготных носителей гена врожденного вывиха бедра в популяции.
Решение.
Генотипы лиц, имеющих врожденный вывих бедра, АА и Аа (доминантное наследование). Здоровые лица имеют генотип аа. Из формулы р2 + 2pq +. q2=1 ясно, что число особей несущих доминантный ген равно (р2+2рq). Однако приведенное в задаче число больных 6/10000 представляет собой лишь одну четвертую (25%) носителей гена А в популяции. Следовательно, р2 + 2pq = (4 х 6)/10 000 = 24/10000. Тогда q2 (число гомозиготных по рецессивному гену особей) равно 1 – (24/10000) = 9976/10000 или 9976 человек.
Ответ: 9976 человек
4.В популяции известны частоты аллелей p = 0,8 и g = 0,2. Определите частоты генотипов.
|
Дано: |
Решение: |
|
|
p = 0,8 |
p2 = 0,64 |
Ответ: частота генотипа АА – 0,64; генотипа аа – 0,04; генотипа Аа – 0,32.
5.Популяция имеет следующий состав: 0,2 АА, 0,3 Аа и 0,50 аа. Найдите частоты аллелей А и а.
|
Дано: |
Решение: |
|
|
p2 = 0,2 |
p = 0,45 |
Ответ: частота аллеля А – 0,45; аллеля а – 0,55.
6.В стаде крупного рогатого скота 49% животных рыжей масти (рецессив) и 51% черной масти (доминанта). Сколько процентов гомо- и гетерозиготных животных в этом стаде?
|
Дано: |
Решение: |
|
|
g2 = 0,49 |
g = 0,7 |
Ответ: гетерозигот 42%; гомозигот по рецессиву – 49%; гомозигот по доминанте – 9%.
7. Вычислите частоты генотипов АА, Аа и аа (в%), если особи аа составляют в популяции 1%.
|
Дано: |
Решение: |
|
|
g2 = 0,01 |
g = 0,1 |
Ответ: в популяции 81% особей с генотипом АА, 18% с генотпом Аа и 1% с генотипом аа.
8. При обследовании популяции каракульских овец было выявлено 729 длинноухих особей (АА), 111 короткоухих (Аа) и 4 безухих (аа). Вычислите наблюдаемые частоты фенотипов, частоты аллелей, ожидаемые частоты генотипов по формуле Харди-Вайнберга.
Это задача по неполному доминированию, поэтому, распределение частот генотипов и фенотипов совпадают и их можно было бы определить, исходя из имеющихся данных.
Для этого надо просто найти сумму всех особей популяции (она равна 844), найти долю длинноухих, короткоухих и безухих сначала в процентах (86.37, 13.15 и 0.47, соответственно) и в долях частот (0.8637, 0.1315 и 0.00474).
Но в задании сказано применить для расчетов генотипов и фенотипов формулу Харди-Вайнберга и, к тому же, рассчитать частоты аллелей генов А и а. Так вот для расчета самих частот аллелей генов без формулы Харди-Вайнберга не обойтись.
Обозначим частоту встречаемости аллеля А во всех гаметах популяции овец буквой р, а частоту встречаемости аллеля а – буквой q. Сумма частот аллельных генов p + q = 1.
Так как по формуле Харди-Вайнберга p2AA + 2pqAa + q2aa = 1 имеем, что частота встречаемости безухих q2 равна 0.00474, то извлекая квадратный корень из числа 0.00474 мы находим частоту встречаемости рецессивного аллеля а. Она равна 0.06884.
Отсюда можем найти частоту встречаемости и доминантного аллеля А. Она равна 1 – 0.06884 = 0.93116.
Теперь по формуле можем вычислить снова частоты встречаемости длинноухих (АА), безухих (аа) и короткоухих (Аа) особей. Длинноухих с генотипом АА будет р2 = 0.931162 = 0.86706, безухих с генотипом аа будет q2 = 0.00474 и короткоухих с генотипом Аа будет 2pq = 0,12820. (Вновь полученные числа, рассчитанные по формуле, почти совпадают с вычисленными изначально, что говорит о справедливости закона Харди-Вайнберга).
ЗАДАЧИ ДЛЯ САМОСТОЯТЕЛЬНОГО РЕШЕНИЯ
1. Одна из форм глюкозурии наследуется как аутосомно-рецессивный признак и встречается с частотой 7:1000000. Определить частоту встречаемости гетерозигот в популяции.
2. Альбинизм общий (молочно-белая окраска кожи, отсутствие меланина в коже, волосяных луковицах и эпителии сетчатки) наследуется как рецессивный аутосомный признак. Заболевание встречается с частотой 1 : 20 000 (К. Штерн, 1965). Определите процент гетерозиготных носителей гена.
3. У кроликов окраска волосяного покрова “шиншилла” (ген Cch) доминирует над альбинизмом (ген Ca). Гетерозиготы CchCa имеют светло-серую окраску. На кролиководческой ферме среди молодняка кроликов шиншилл появились альбиносы. Из 5400 крольчат 17 оказались альбиносами. Пользуясь формулой Харди-Вайнберга, определите, сколько было получено гомозиготных крольчат с окраской шиншилла.
4. Популяция европейцев по системе групп крови резус содержит 85% резус положительных индивидуумов. Определите насыщенность популяции рецессивным аллелем.
5. Подагра встречается у 2% людей и обусловлена аутосомным доминантным геном. У женщин ген подагры не проявляется, у мужчин пенетрантность его равна 20% (В.П. Эфроимсон, 1968). Определите генетическую структуру популяции по анализируемому признаку, исходя из этих данных.
Решение 1. Обозначим аллельный ген, отвечающий за проявление глюкозурии а, так как сказано, что это заболевание наследуется как рецессивный признак. Тогда аллельный ему доминантный ген, отвечающий за отсутствие болезни обозначим А.
Здоровые особи в популяции людей имеют генотипы АА и Аа; больные особи имеют генотип только аа.
Обозначим частоту встречаемости рецессивного аллеля а буквой q, а доминантного аллеля А – буквой р.
Поскольку нам известно, что частота встречаемости больных людей с генотипом аа (а это значит q2) равна 0,000007, то q = 0,00264575
Так как p + q = 1, то р = 1 — q = 0,9973543, и p2 = 0,9947155
Теперь подставив значения р и q в формулу: p2AA + 2pqAa + q2aa = 1,
найдем частоту встречаемости гетерозиготных особей 2pq в популяции людей: 2pq = 1 – p2 — q2 = 1 – 0,9947155 – 0,000007 = 0,0052775.
Решение 2. Так как этот признак рецессивный, то больные организмы будут иметь генотип аа — это их частота равна 1 : 20 000 или 0,00005.
Частота аллеля а составит корень квадратный из этого числа, то есть 0,0071. Частота аллеля А составит 1 — 0,0071 = 0,9929, а частота здоровых гомозигот АА будет 0,9859. Частота всех гетерозигот 2Аа = 1 — (АА + аа) = 0,014 или 1,4%.
Решение 3. Примем 5400 штук всех кроликов за 100%, тогда 5383 кролика (сумма генотипов АА и Аа) составит 99,685% или в частях это будет 0,99685.
q2 + 2q(1 – q) = 0,99685 – это частота встречаемости всех шиншилл и гомозиготных (АА), и гетерозиготных (Аа).
Тогда из уравнения Харди-Вайнберга: q2 AA+ 2q(1 – q)Aa + (1 – q)2aa = 1 , находим (1 – q)2 = 1 – 0,99685 = 0,00315 – это частота встречаемости альбиносных кроликов с генотипом аа. Находим чему равна величина 1 – q. Это корень квадратный из 0,00315 = 0,056. А q тогда равняется 0,944.
q2 равняется 0,891, а это и есть доля гомозиготных шиншил с генотипом АА. Так как эта величина в % составит 89,1% от 5400 особей, то количество гомозиготных шиншилл будет 4811 шт.
Решение 4. Нам известно, что аллельный ген, отвечающий за проявление резус положительной крови является доминантным R (обозначим частоту его встречаемости буквой p), а резус отрицательный – рецессивным r (обозначим частоту встречаемости его буквой q).
Поскольку в задаче сказано, что на долю p2RR + 2pqRr приходится 85% людей, значит на долю резус-отрицательных фенотипов q2rr будет приходиться 15% или частота встречаемости их составит 0,15 от всех людей европейской популяции.
Тогда частота встречаемости аллеля r или ”насыщенность популяции рецессивным аллелем” (обозначенная буквой q) составит корень квадратный из 0,15 = 0,39 или 39%.
Решение 5. Подагра встречается у 2% людей и обусловлена аутосомным доминантным геном. У женщин ген подагры не проявляется, у мужчин пенетрантность его равна 20% (В.П. Эфроимсон, 1968). Определите генетическую структуру популяции по анализируемому признаку, исходя из этих данных.
Так как подагра выявляется у 2% мужчин, то есть у 2 человек из 100 с пенетрантностью 20%, то реально носителями генов подагры является в 5 раз больше мужчин, то есть 10 человек из 100.
Но, так как мужчины составляют лишь пол популяции, то всего людей с генотипами АА + 2Аа в популяции будет 5 человек из 100, а, значит, 95 из 100 будут с генотипом аа.
Если частота встречаемости организмов с генотипами аа составляет 0,95, то частота встречаемости рецессивного аллеля а в этой популяции равна корню квадратному из числа 0,95 = 0,975. Тогда частота встречаемости доминантного аллеля ”А” в этой популяции равна 1 – 0,975 = 0,005.
При определённых условиях популяция находится в состоянии генетического равновесия, т. е. её генофонд не изменяется из поколения в поколение. Это принцип равновесия, или закон Харди — Вайнберга.
В идеальной популяции в ряду поколений сохраняется без изменений постоянное соотношение частот генов, гомозигот и гетерозигот.
Идеальная популяция характеризуется следующими признаками:
- число особей достаточно большое;
- особи свободно скрещиваются;
- не происходят мутации;
- нет миграции из соседних популяций;
- отсутствует естественный отбор.
Закон Харди — Вайнберга позволяет определять частоты генов и генотипов.
Частоту доминантного гена
A
обычно обозначают буквой
p
, а частоту рецессивного гена
a
— буквой
q
.
Составим схему скрещивания и установим возможные сочетания аллелей гена и их частоты.
|
Аллель (частота) |
A (p) | a (q) |
| A (p) | AA (p2) | Aa (pq) |
| a (q) | Aa (pq) | aa (q2) |
Значит, частота доминантных гомозигот
AA
равна
p2
, частота гетерозигот
Aa
—
2pq
, а частота рецессивных гомозигот
aa
—
q2
.
Если аллельных генов два, то сумма их частот равна единице (или (100) %):
Сумма частот генотипов тоже равна единице (или (100) %):
По формуле Харди — Вайнберга можно определять частоты генов в природных популяциях, например вычислять частоты полезных и вредных мутаций в популяциях растений и животных при восстановлении исчезающих видов или создании новых сортов и пород.
В естественных условиях идеальных популяций не существует. Мутации происходят всегда, имеют место миграции особей и отбор. Но для количественной оценки многих генетических явлений закон Харди — Вайнберга применим.
Популяционная генетика – это раздел генетики, изучающий генетическое строение и динамику генетического состава популяций – группы организмов, принадлежащих к одному и тому же виду и занимающих ограниченную географическую область обитания. Именно этот раздел науки является основой синтетической теории эволюции, принятой в настоящее время большинством учёных.
Генотип совместно с внешней средой определяет фенотип организма. Фенотипы, приспособленные к условиям места обитания, сохраняются в процессе естественного отбора; неприспособленные фенотипы подавляются. Для популяции в целом судьба отдельной особи значения не имеет.
Генетическое разнообразие популяций на примере цветков различной окраски
Генофонд – это многообразие генов и аллелей, имеющихся в популяции. В каждой популяции, размножающейся половым путём, генофонд постоянно изменяется; популяция эволюционирует. Новые сочетания генов проходят непрерывный естественный отбор, определяющий в конечном итоге, какие гены будут переданы следующим поколениям.
Частотой аллеля называют отношение количества данных аллелей у всех особей к общему количеству аллелей в популяции. Частоту доминантного аллеля обычно обозначают буквой p, частоту рецессивного аллеля – буквой q. Если ген представлен двумя аллелями, то выполняется математическое равенство
p + q = 1.
Таким образом, зная частоту одного из аллелей, можно определить частоту и другого аллеля. Так, если частота доминантного аллеля равна 78 %, то частота рецессивного аллеля равняется q = 1 – p = 1 – 0,78 = 0,22 (или 22 %).
Важным понятием популяционной генетики является частота аллеля – его доля среди имеющих аллелей.
