Время на прочтение
10 мин
Количество просмотров 23K

Знай своего врага ― одна из максим, которой руководствуются специалисты по информационной безопасности. Она касается и зловредов. Существуют сотни инструментов, которые помогают исследовать вредоносное ПО. К счастью, многие из них бесплатны и имеют открытый исходный код.
Под катом мы собрали онлайн-сканеры подозрительных файлов, некоторые инструменты для статического и динамического анализа, системы для описания и классификации угроз и, конечно, репозитории с малварью, которую можно исследовать.
Это неполный перечень ПО, которое используют исследователи от мира информационной безопасности. Так, для расследования уже совершенных атак, применяют иной арсенал.
Деление на группы в статье не претендует на звание классификации, оно сделано для удобства чтения и во многом условно. Отдельные инструменты могли попасть сразу в несколько групп и оказались в конкретных разделах благодаря грубому авторскому произволу.
Онлайн-службы для анализа зловредов
-
VirusTotal ― бесплатная служба для анализа подозрительных файлов и ссылок. В представлении не нуждается.
-
Intezer ― детектор зловредов, поддерживающий динамический и статический анализ.
-
Triage ― онлайн-сервис для анализа больших объемов вредоносного ПО с функцией статического анализа образцов.
-
FileScan.IO — служба анализа вредоносных программ, с динамическим анализом и функцией извлечения индикаторов компрометации (IOC).
-
Sandbox.pikker ― онлайн-версия известной системы анализа вредоносных программ Cuckoo Sandbox. Предоставляет подробный отчет с описанием поведения файла при выполнении в реалистичной, но изолированной среде далеко в облаке.
-
Manalyzer ― бесплатный сервис, для статического анализа PE-файлов и выявления маркеров нежелательного поведения. Имеет офлайн-версию.
-
Opswat ― сканирует файлы, домены, IP-адреса и хеши при помощи технологии Content Disarm & Reconstruction.
-
InQuest Labs ― сервис для сканирования текстовых документов файлов Microsoft и Open Office, электронных таблиц и презентаций. Работает на базе механизмов Deep File Inspection (DFI).
-
Any Run ― еще одна онлайн-песочница с красивым интерфейсом и дополнительными опциями.
-
Yoroi ― итальянская служба анализа подозрительных файлов на базе песочницы. Переваривает PE (например, .exe-файлы), документы (doc и PDF), файлы сценариев (типа wscript, Visual Basic) и APK, но мучительно медленно готовит отчеты.
-
Unpacme ― онлайн-сервис для автоматической распаковки вредоносных программ и извлечения артефактов.
-
Malwareconfig ― веб-приложение для извлечения, декодирования и отображения параметров конфигурации распространенных вредоносных программ.
-
Malsub ― фреймворк Python RESTful для работы с API онлайн-сервисов для анализа вредоносного ПО.
Дистрибутивы для анализа вредоносных программ
-
REMnux — готовый дистрибутив реверс-инжиниринга и анализа вредоносных программ на основе Ubuntu. Предоставляет полный пакет необходимых утилит с открытым исходным кодом и включает многие инструменты, упомянутые ниже.
-
Tsurugi Linux ― Linux-дистрибутив, ориентированный на цифровую криминалистику, анализ вредоносных программ и разведку по открытым источникам.
-
Flare-vm ― настраиваемый дистрибутив для анализа вредоносных программ на базе Windows.
Комбайны и тулкиты
-
Ghidra Software Reverse Engineering Framework ― продвинутая среда для реверс-инжиниринга ПО под Windows, macOS и Linux. Поддерживает широкий набор инструкций процессора и исполняемых форматов, работает и в ручном, и в автоматическом режимах, поддерживает пользовательские сценарии. Некоторые такие заготовки, упрощающие анализ вредоносного ПО, можно найти в репозитории Ghidra-Scripts.
-
FLARE VM ― настраиваемый дистрибутив для анализа вредоносных программ на базе Windows 10.
-
Indetectables Toolkit ― набор из 75 приложений для реверс-инжиниринга и анализа двоичных файлов и вредоносных программ.
-
MobSF ― платформа для автоматизированного статического и динамического анализа и оценки безопасности мобильных приложений.
-
CyberChef — приложение для анализа и декодирования данных без необходимости иметь дело со сложными инструментами или языками программирования.
-
Pev ― набор инструментов для анализа подозрительных PE-файлов. Работает в Windows, Linux и macOS.
-
Vivisect ― фреймворк Python, который включает функции дизассемблера, статического анализатора, символьного выполнения и отладки.
-
Pharos ― экспериментальный фреймворк, объединяющий шесть инструментов для автоматизированного статического анализа бинарных файлов.
Инструменты статического анализа и реверс-инжиринга
-
IDA ― дизассемблер и отладчик для Windows с бесплатной версией и возможность расширения функциональности при помощи open-source-плагинов, например, Diaphora и FindYara.
-
Binary ninja — популярная альтернатива IDA с бесплатной облачной версией и частично открытым исходным кодом.
-
ret-sync ― набор плагинов, которые позволяют синхронизировать сеансы в дизассемблерах IDA, Ghidra и Binary Ninja.
-
Binary Analysis Platform ― мультиплатформенный набор утилит и библиотек для анализа машинного кода. Поддерживает x86, x86-64, ARM, MIPS, PowerPC. Другие архитектуры могут быть добавлены в виде плагинов.
-
Radare2 ― UNIX-подобная среда для реверс-инжиниринга. Позволяет анализировать, эмулировать, отлаживать, модифицировать и дизассемблировать любой бинарник. Может работать через удаленный сервер gdb.
-
Rizin ― форк Radeone2 с упором на удобство использования и стабильность. Включает все необходимое для полноценного бинарного анализа.
-
cutter ― бесплатная платформа для реверс-инжиниринга с открытым исходным кодом и плагином для интеграции с декомпилятором Ghidra. Построена на базе rizin. Предоставляет удобный пользовательский интерфейс.
-
Un{i}packer ― кроссплатформенная альтернатива деббагеру x64Dbg.
-
diStorm ― быстрый дизассемблер для анализа вредоносного шелл-кода.
-
angr ― мультиплатформенный фреймворк для исследования бинарных файлов.
-
Binwalk ― простой в использовании инструмент для анализа, извлечения образов и реверс-инжиниринга микропрограмм.
-
4n4lDetector ― инструмент для быстрого анализа исполняемых файлов Windows. Показывает PE-заголовок и его структуру, содержимое разделов, различные типы строк. Облегчает идентификацию вредоносного кода внутри анализируемых файлов. Исходный код не опубликован.
-
capa ― анализирует возможности исполняемых файлов. Вы скармливаете PE, ELF и шеллкод, а capa сообщает, что, может сделать программа.
-
LIEF ― кроссплатформенная библиотека для анализа, изменения и абстрагирования форматов ELF, PE, MachO, DEX, OAT, ART и VDEX без использования дизассемблера.
-
IDR ― декомпилятор исполняемых файлов (EXE) и динамических библиотек (DLL), написанный на Delphi. Часто используется для анализа банковских вредоносных программ.
-
Manalyze ― синтаксический анализатор PE-файлов с поддержкой плагинов. Лежит в основе вышеупомянутого онлайн-сервиса Manalyzer.
-
PortEx ― библиотека Java для статического анализа переносимых исполняемых файлов с акцентом на анализ вредоносных программ и устойчивость к искажениям PE.
-
PE-bear ― анализатор файлов PE-формата со встроенным Hex-редактором и возможностью детектирования различных упаковщиков по сигнатурам.
-
CFF Explorer ― аналог PE-bear, созданный Эриком Пистелли. Представляет собой набор инструментов: шестнадцатеричный редактор, сканер и менеджер подписей, дизассемблер, обходчик зависимостей и т. д. Первый PE-редактор с поддержкой внутренних структур .NET.
-
Qu1cksc0pe ― универсальный инструмент для статического анализа исполняемых файлов Windows, Linux, OSX, а также APK.
-
Quark-Engine ― система оценки вредоносного ПО для Android, игнорирующая обфускацию. Входит в комплект поставки Kali Linux и BlackArch.
-
Argus-SAF ― платформа для статического анализа Android-приложений.
-
RiskInDroid ― инструмент для оценки потенциальной опасности Android-приложений на основе методов машинного обучения.
-
MobileAudit ― SAST-анализатор вредоносных программ для Android.
-
Obfuscation Detection ― набор скриптов для автоматического обнаружения запутанного кода.
-
de4dot ― деобфускатор и распаковщик .NET, написанный на C#.
-
FLARE ― статический анализатор для автоматической деобфускации бинарных файлов вредоносных программ.
-
JSDetox ― утилита для анализа вредоносного Javascript-кода с функциями статического анализа и деобфускации.
-
XLMMacroDeobfuscator ― используется для декодирования запутанных макросов XLM. Использует внутренний эмулятор XLM для интерпретации макросов без полного выполнения кода. Поддерживает форматы xls, xlsm и xlsb.
Инструменты динамического анализа и песочницы
-
Cuckoo Sandbox — платформа для анализа разнообразных вредоносных файлов в виртуализированных средах Windows, Linux, macOS и Android. Поддерживает YARA и анализ памяти зараженной виртуальной системы при помощи Volatility.
-
CAPE ― песочница с функцией автоматической распаковки вредоносного ПО. Построена на базе Cuckoo. Позволяет проводить классификацию на основе сигнатур Yara.
-
Qubes OS ― защищенная операционная система, основанная на концепции разграничения сред. Использует гипервизор Xen и состоит из набора отдельных виртуальных машин. Может служить площадкой для исследования вредоносов, однако не включает в себя специализированные инструменты для их анализа.
-
x64dbg ― отладчик с открытым исходным кодом под Windows, предназначенный для реверс-инжиниринга вредоносных файлов. Выделяется обширным списком специализированных плагинов.
-
DRAKVUF ― безагентная система бинарного анализа типа «черный ящик». Позволяет отслеживать выполнение бинарных файлов без установки специального программного обеспечения на виртуальной машине. Требует для работы процессор Intel с поддержкой виртуализации (VT-x) и Extended Page Tables (EPT).
-
Noriben ― простая портативная песочница для автоматического сбора и анализа информации о вредоносных программах и составления отчетов об их действиях.
-
Process Hacker ― продвинутый диспетчер задач, позволяющий наблюдать за поведением подозрительных программ в реальном времени. Отображает сетевые подключения, активные процессы, информацию об использовании памяти и обращениях к жестким дискам, использовании сети. Также отображает трассировки стека.
-
Sysmon ― системная служба Windows, регистрирующая активность системы в журнале событий Windows. Предоставляет подробную информацию о происходящем в системе: о создании процессов, сетевых подключениях времени создания и изменения файлов. Работает с заранее созданными конфигурационными файлами, которые можно найти на github.
-
hollows_hunter ― сканер процессов, нацеленный на обнаружение вредоносных программ и сбор материалов для дальнейшего анализа. Распознает и создает дамп различных имплантов в рамках сканируемых процессов. Основан на PE-sieve.
-
Sandboxapi ― API Python для создания интеграций с различными проприетарными песочницами.
-
DECAF ― Dynamic Executable Code Analysis Framework ― платформа для анализа бинарных файлов, основанная на QEMU. Поддерживает 32-разрядные версии Windows XP/Windows 7/linux и X86/arm.
-
Frida ― внедряет фрагменты JavaScript в нативные приложения для Windows, Mac, Linux, iOS и Android, что позволяет изменять и отлаживать запущенные процессы. Frida не требует доступа к исходному коду и может использоваться на устройствах iOS и Android, которые не взломаны или не имеют root-доступа.
-
box-js ― инструмент для изучения вредоносного JavaScript-ПО с поддержкой JScript/WScript и эмуляцией ActiveX.
-
Fibratus ― инструмент для исследования и трассировка ядра Windows.
-
analysis-tools ― большая коллекция инструментов динамического анализа для различных языков программирования.
Инструменты анализа сетевой активности
-
MiTMProxy ― перехватывающий HTTP-прокси с поддержкой SSL/TLS.
-
Wireshark ― известный универсальный инструмент для анализа сетевого трафика.
-
Zeek ― фреймворк для анализа трафика и мониторинга безопасности, ведет подробные журналы сетевой активности.
-
Maltrail ― система обнаружения вредоносного трафика на базе общедоступных черных списков сетевых адресов.
-
PacketTotal ― онлайн-движок для анализа .pcap-файлов и визуализации сетевого трафика.
Хранение и классификация и мониторинг
-
YARA ― инструмент, предназначенный для помощи в идентификации и классификации зловредов. Позволяет создавать описания семейств вредоносных программ в виде правил ― в общепринятом удобном для обработки формате. Работает на Windows, Linux и Mac OS X.
-
Yara-Rules ― обширный список правил для YARA, позволяет быстро подготовить этот инструмент к использованию.
-
yarGen ― генератор правил YARA из подозрительных фрагментов кода, найденных в файлах вредоносных программ. Автоматизирует процесс создания новых правил.
-
Awesome YARA ― пополняемая коллекция инструментов, правил, сигнатур и полезных ресурсов для пользователей YARA.
-
YETI ― платформа, для сбора и агрегации данных об угрозах в едином репозитории. Умеет обогащать загруженные данные, снабжена удобным графическим интерфейсом и развитым API.
-
Viper ― утилита для организации, анализа и менеджмента личной коллекции вредоносных программ, образцов эксплойтов и скриптов. Хорошо работает в связке с Cuckoo и YARA.
-
MISP ― ПО для сбора, хранения обмена индикаторами компрометации и данными, полученными в результате анализа вредоносных программ.
-
Malice ― фреймворк для анализа вредоносных программ, который называют open source версией VirusTotal.
-
Hostintel ― модульное приложение на Python для сбора сведений о вредоносных хостах.
-
mailchecker — проверка электронной почты на разных языках. Утилита снабжена базой данных из более чем 38 000 поддельных провайдеров электронной почты.
-
URLhaus ― портал для обмена URL-адресами, которые используются для распространения вредоносных программ.
-
All Cybercrime IP Feeds ― агрегатор опасных IP-адресов с акцентом на хакерские атаки, распространение вредоносного ПО и мошенничество. Использует данные из более чем 300 источников.
-
Machinae — инструмент для сбора информации о зловредах из общедоступных источников. Позволяет получить списки подозрительных IP-адресов, доменных имен, URL, адресов электронной почты, хешей файлов и SSL fingerprints.
-
ThreatIngestor — конвейер для сбора данных об угрозах из Twitter, GitHub, RSS каналов и других общедоступных источников.
-
InQuest REPdb и InQuest IOCdb агрегаторы IOC из различных открытых источников.
-
IntelOwl — решение для получения данных при помощи единого API. Предоставляет централизованный доступ к ряду онлайн-анализаторов вредоносных программ.
-
Malwoverview ― инструмент для классификации и сортировки вредоносных программ IOC и хешей с возможностью генерации отчетов по результатам статического и динамического анализа.
-
dnstwist ― инструмент для поиска мошеннических доменов, маскирующихся под ваш сайт.
Образцы вредоносных программ
В этом разделе собраны ссылки на опасные вредоносные программы. Они не деактивированы и представляют реальную угрозу. Не скачивайте их, если не уверены в том, что делаете. Используйте для исследований только изолированные программные среды. В этих коллекциях много червей, которые заразят все, до чего смогут дотянуться.
Ссылки предоставляются исключительно в научных и образовательных целях.
-
theZoo ― активно пополняемая коллекция вредоносных программ, собранная специально для исследователей и специалистов по кибербезопасности.
-
Malware-Feed ― коллекция вредоносных программ от virussamples.com.
-
Malware samples ― пополняемый github-репозиторий с вредоносами, к которым прилагаются упражнения по анализу вредоносного ПО.
-
vx-underground ― еще одна обширная и постоянно растущая коллекция бесплатных образцов вредоносных программ.
-
Malshare ― репозиторий, который помимо образцов малвари предлагает дополнительные данные для YARA.
-
MalwareBazaar ― проект, целью которого является обмен образцами вредоносного ПО с поставщиками антивирусных программ и сообществом экспертов по информационной безопасности. Поддерживает собственное API.
-
Virusbay ― онлайн-платформа для совместной работы, которая связывает специалистов из центров мониторинга информационной безопасности с исследователями вредоносных программ. Представляет собой специализированную социальную сеть.
-
VirusShare ― большой репозиторий вредоносных программ с обязательной регистрацией.
-
The Malware Museum ― вирусы, распространявшиеся в 1980-х и 1990-х годах. Практически безобидны на фоне остальных ссылок из этого раздела и представляют скорее историческую ценность.
Многие современные зловреды выходят из-под пера талантливых профессиональных программистов. В них используются оригинальные пути проникновения в системные области данных, новейшие уязвимости и ошибки в безопасности операционных систем и целые россыпи грязных хаков. Если проявить осторожность, ум и внимательность, во вредоносном ПО можно найти немало интересного.
Удачной охоты!
Запрос «Вирус» перенаправляется сюда; см. также другие значения.
| Вирусы | |||||||
|---|---|---|---|---|---|---|---|
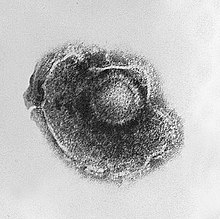
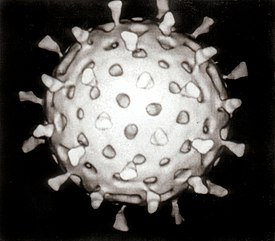 Ротавирус, компьютерная реконструкция на основе данных электронной криомикроскопии |
|||||||
| Научная классификация | |||||||
|
Группа: Вирусы |
|||||||
| Международное научное название | |||||||
| Viruses [комм. 1] | |||||||
| Реалмы[комм. 2] | |||||||
систематика 1 класса (Naldaviricetes) и 15 семейств вирусов не определена (как и 2 семейств вироидов и 2 семейств сателлитов), они не входят в описанные реалмы[1] |
|||||||
|
Ви́рус (лат. virus) — неклеточный инфекционный агент, который может воспроизводиться только внутри клеток[комм. 3]. Вирусы поражают все типы организмов, от растений и животных до бактерий и архей[2] (вирусы бактерий обычно называют бактериофагами). Обнаружены также вирусы, способные реплицироваться только в присутствии других вирусов (вирусы-сателлиты).
Со времени публикации в 1892 году статьи Дмитрия Ивановского, описывающей небактериальный патоген растений табака[3][4], и открытия в 1898 году Мартином Бейеринком вируса табачной мозаики[5] были детально описаны более 6 тысяч видов вирусов[6], хотя предполагают, что их существует более ста миллионов[7]. Вирусы обнаружены почти в каждой экосистеме на Земле, они являются самой многочисленной биологической формой[8][9]. Изучением вирусов занимается наука вирусология, раздел микробиологии.
Геномы вирусов могут быть представлены как ДНК, так и РНК, причем в обоих случаях как одноцепочечной, так и двуцепочечной. Некоторые вирусы способны к обратной транскрипции. Кроме того, у некоторых РНК-содержащих вирусов в репликации участвует рибозим, что сближает их с вироидами. Однако все вирусы, в отличие от вироидов, образуют белковые капсиды, в которые заключён их генетический материал.
У животных вирусные инфекции вызывают иммунный ответ, который чаще всего приводит к уничтожению болезнетворного вируса. Иммунный ответ также можно вызвать вакцинами, дающими активный приобретённый иммунитет против конкретной вирусной инфекции. Однако некоторым вирусам, в том числе вирусу иммунодефицита человека и возбудителям вирусных гепатитов, удаётся избежать иммунного ответа, вызывая хроническую болезнь. Антибиотики не действуют на вирусы, однако было разработано несколько противовирусных препаратов.
Термин «вирус» является общим для обозначения инфекционного агента, способного паразитировать внутри клеток. Для обозначения единичной стабильной вирусной частицы, покинувшей клетку и способной инфицировать другие клетки того же типа, применяется термин «вирион»[10].
Этимология названия[править | править код]
Слово «вирус» образовано от лат. virus — «болезнетворный яд», «слизь»[11]. Для обозначения инфекционного заболевания оно впервые было применено в 1728 году, в 1790-х годах использовалось для обозначения агента, способного вызывать инфекционное заболевание[12]. Для обозначения субмикроскопического инфекционного агента латинское слово virus начало использоваться с работы голландского микробиолога Мартина Бейеринка, опубликованной в 1898 году, однако на тот момент вирус был ошибочно принят за жидкость, которая проходила через фильтр, не пропускающий бактерии[13]. Сам вирус был открыт в 1892 году Дмитрием Ивановским[4]. В подобных случаях инфекционного агента стали называть «фильтрующимся вирусом», чтобы отличать его от бактерий. Хотя таких фильтрующихся вирусов к 1920-м годам было известно уже достаточно, их природа была всё ещё непонятна, а в 1930-х годах этот термин перестали использовать в пользу более простого слова «вирус» для обозначения небактериальных агентов. В конце 1930-х годов впервые удалось визуализировать вирусы с помощью электронного микроскопа и их природа, наконец, стала понятна[14]. Первое использование производного от virus термина «вирион» для обозначения отдельных вирусных частиц датируется 1959 годом[15].
История исследований[править | править код]

По мере накопления данных об инфекционных заболеваниях различных организмов стало очевидно, что далеко не все из них вызываются патогенами, известными на тот момент — бактериями, протистами или микроскопическими грибками. В частности, Луи Пастер не смог найти агент, вызывающий бешенство, и предполагал, что этот патоген слишком мал, чтобы увидеть его в микроскоп[16]. В 1884 году французский микробиолог Шарль Шамберлан изобрёл фильтр (сейчас известный как фильтр Шамберлана или фильтр Шамберлана—Пастера), поры которого меньше бактерий. С помощью этого фильтра можно полностью удалить бактерии из раствора[17]. В 1892 году русский биолог Дмитрий Ивановский использовал его для изучения вида, сейчас известного как вирус табачной мозаики. Его эксперименты показали, что экстракт перетёртых листьев заражённых растений табака сохраняет инфекционные свойства и после фильтрации. Ивановский предположил, что инфекция может вызываться токсином, выделяемым бактериями, однако он не развил эту идею[18]. В то время считалось, что любой инфекционный агент можно выделить на фильтре и выращивать в питательной среде — таков один из постулатов микробной теории болезней[5]. Кроме того, Ивановский в оптическом микроскопе наблюдал в заражённых клетках растений кристаллоподобные тела, которые в современном понимании являлись скоплениями вирусов, впоследствии они были названы «кристаллами Ивановского»[19]. В 1898 году голландский микробиолог Мартин Бейеринк повторил эксперименты Ивановского и пришёл к выводу, что прошедший сквозь фильтр инфекционный материал есть не что иное, как новая форма инфекционных агентов[20]. Он подметил, что агент размножался только в делящихся клетках, однако его опыты не выявили того, что он представляет собой частицы. Бейеринк назвал его Contagium vivum fluidum (дословно лат. растворимый живой микроб) и вновь ввёл в употребление слово «вирус»[18]. Он утверждал, что по своей природе вирус жидкий. Эта теория впоследствии была опровергнута Уэнделлом Стэнли, доказавшим, что вирусы представляют собой частицы[18]. В том же году Фридрих Лёффлер и Пауль Фрош обнаружили первый вирус животных — возбудитель ящура (афтовирус), пропустив его через схожий фильтр[21].
В начале XX века английский бактериолог Фредерик Туорт открыл группу вирусов, инфицирующих бактерии (сейчас они известны как бактериофаги[22] или просто фаги), а франко-канадский микробиолог Феликс д’Эрелль описал вирусы, которые при добавлении к бактериям на агаре образуют вокруг себя пространство с мёртвыми бактериями. Д’Эрелль сделал точные разведения суспензии этих вирусов и установил наибольшее разведение (наименьшую концентрацию вирусов), при котором не все бактерии погибают, но тем не менее образуются отдельные области с мёртвыми клетками. Подсчитав число таких областей и учитывая коэффициент разведения, он определил число вирусных частиц в первоначальной суспензии[23]. Фаги были объявлены потенциальным средством от болезней, таких как тиф и холера, однако это было забыто в связи с открытием уникальных свойств пенициллина. Изучение фагов дало информацию о явлении «включения» и «выключения» генов, а также позволило использовать их для введения в бактериальный геном чужеродных генов.
К концу XIX века было известно, что вирусы обладают инфекционными свойствами, способны проходить через фильтры и нуждаются в живом хозяине для размножения. В то время вирусы в исследовательских целях культивировали только в растениях и животных. В 1906 году Росс Грэнвилл Гаррисон изобрёл метод выращивания тканей в лимфе, и в 1913 году Штейнард, Израэли и Ламберт использовали этот метод при выращивании вируса осповакцины на фрагментах ткани роговицы морских свинок[24]. В 1928 году Г. Б. Мэйтланд и М. К. Мэйтланд вырастили вирус осповакцины на суспензии из измельчённых куриных почек. Этот метод не применялся широко до конца 1950-х годов, когда в больших масштабах стали выращивать полиовирус для производства вакцины[25].
Другое крупное достижение принадлежит американскому патологу Эрнесту Уильяму Гудпасчеру; в 1939 г он вырастил вирус гриппа и несколько других вирусов в оплодотворённых куриных яйцах[26]. В 1949 году Джон Франклин Эндерс, Томас Уэллер и Фредерик Роббинс вырастили полиовирус на культуре клеток зародыша человека. Это был первый вирус, выращенный не на тканях животных или яйцах. Эта работа дала возможность Джонасу Солку создать эффективную полиовакцину (вакцину против полиомиелита)[27].
Первые изображения вирусов были получены после изобретения электронного микроскопа немецкими инженерами Эрнстом Руской и Максом Кноллем[28]. В 1935 году американский биохимик и вирусолог Уэнделл Мередит Стэнли тщательно изучил вирус табачной мозаики и обнаружил, что он по большей части состоит из белка[29]. Спустя короткое время этот вирус был разделён на белковую и РНК-составляющую[30]. Вирус табачной мозаики был кристаллизован первым среди вирусов, что позволило многое узнать о его структуре. Первая рентгенограмма кристаллизованного вируса была получена Берналем и Фэнкухеном в конце 1930-х годов. На основании полученных ею изображений Розалинд Франклин в 1955 году определила полную структуру вируса[31]. В том же году Хайнц Френкель-Конрат и Робли Уилльямс показали, что очищенная РНК вируса табачной мозаики и белок оболочки способны к самосборке в функциональный вирус. Это позволило им предположить, что подобный механизм лежит в основе сборки вирусов внутри клеток-хозяев[32].
Вторая половина XX века стала периодом расцвета вирусологии. В то время было открыто свыше 2000 видов вирусов животных, растений и бактерий[33]. В 1957 году были открыты лошадиный артеривирус и возбудитель вирусной диареи коров (пестивирус). В 1963 году Барух Бламберг открыл вирус гепатита B[34], а в 1965 году Хоуард Темин описал первый ретровирус. В 1970 году Темин и Дейвид Балтимор независимо друг от друга описали обратную транскриптазу, ключевой фермент, с помощью которого ретровирусы синтезируют ДНК-копии своих РНК[35]. В 1983 году группа учёных во главе с Люком Монтанье из Института Пастера во Франции впервые выделила ретровирус, известный сейчас как ВИЧ[36].
В 2002 году в Нью-Йоркском университете был создан первый синтетический вирус (вирус полиомиелита)[37].
Происхождение[править | править код]
Появление вирусов на эволюционном древе жизни неясно: некоторые из них могли образоваться из плазмид, небольших молекул ДНК, способных передаваться от одной клетки к другой, в то время как другие могли произойти от бактерий. В эволюции вирусы являются важным звеном горизонтального переноса генов, обуславливающего генетическое разнообразие[38]. Некоторые учёные считают вирусы особой формой жизни, так как они имеют генетический материал, способны создавать себе подобные вирусы, и эволюционируют путем естественного отбора. Однако у вирусов отсутствуют важные характеристики (такие как клеточное строение и собственный обмен веществ), без которых их нельзя отнести к живому. Так как они обладают некоторыми, но не всеми свойствами живого, вирусы описываются как «организмы на краю жизни».
Вирусы найдены везде, где есть жизнь, и, вероятно, вирусы существуют с момента появления первых живых клеток[39]. Происхождение вирусов неясно, поскольку они не оставляют каких бы то ни было ископаемых остатков, а их родственные связи можно изучать только методами молекулярной филогенетики[40].
Гипотезы о происхождении вирусов[править | править код]
Существует три основные гипотезы происхождения вирусов: регрессивная гипотеза, гипотеза клеточного происхождения и гипотеза коэволюции[41][42].
Регрессивная гипотеза[править | править код]
Согласно этой гипотезе, вирусы когда-то были мелкими клетками, паразитирующими в более крупных клетках. С течением времени эти клетки предположительно утратили гены, которые были «лишними» при паразитическом образе жизни. Эта гипотеза основывается на наблюдении, что некоторые бактерии, а именно риккетсии и хламидии, представляют собой клеточные организмы, которые тем не менее подобно вирусам могут размножаться только внутри другой клетки. Эту гипотезу также называют гипотезой дегенерации[43][44] или гипотезой редукции[45].
Гипотеза клеточного происхождения[править | править код]
Некоторые вирусы могли появиться из фрагментов ДНК или РНК, которые «высвободились» из генома более крупного организма. Такие фрагменты могут происходить от плазмид (молекул ДНК, способных передаваться от клетки к клетке) или от транспозонов (молекул ДНК, реплицирующихся и перемещающихся с места на место внутри генома)[46]. Транспозоны, которые раньше называли «прыгающими генами», являются примерами мобильных генетических элементов, возможно, от них могли произойти некоторые вирусы. Транспозоны были открыты Барбарой Мак-Клинток в 1950 году в кукурузе[47]. Эту гипотезу также называют гипотезой кочевания[5][48] или гипотезой побега[45].
Гипотеза коэволюции[править | править код]
Эта гипотеза предполагает, что вирусы возникли из сложных комплексов белков и нуклеиновых кислот в то же время, что и первые на Земле живые клетки, и зависят от клеточной жизни вот уже миллиарды лет. Помимо вирусов, существуют и другие неклеточные формы жизни. Например, вироиды — это молекулы РНК, которые не рассматриваются как вирусы, потому что у них нет белковой оболочки. Тем не менее, ряд характеристик сближает их с некоторыми вирусами, а потому их относят к субвирусным частицам[49]. Вироиды являются важными патогенами растений[50]. Они не кодируют собственные белки, однако взаимодействуют с клеткой-хозяином и используют её для осуществления репликации своей РНК[51]. Вирус гепатита D имеет РНК-геном, схожий с геномом вироидов, однако сам не способен синтезировать белок оболочки. Для формирования вирусных частиц он использует белок капсида вируса гепатита B и может размножаться только в клетках, заражённых этим вирусом. Таким образом, вирус гепатита D является дефектным вирусом[52]. Вирофаг Спутник схожим образом зависит от мимивируса, поражающего простейшее Acanthamoeba castellanii[53]. Эти вирусы зависят от присутствия в клетке-хозяине другого вируса и называются вирусами-сателлитами. Подобные вирусы демонстрируют, как может выглядеть промежуточное звено между вирусами и вироидами[54][55].
Каждая из этих гипотез имеет свои слабые места: регрессивная гипотеза не объясняет, почему даже мельчайшие клеточные паразиты никак не походят на вирусы. Гипотеза побега не даёт объяснения появлению капсида и других компонентов вирусной частицы. Гипотеза коэволюции противоречит определению вирусов как неклеточных частиц, зависимых от клеток-хозяев[45].
Тем не менее, в настоящее время многие специалисты признают вирусы древними организмами, появившимися, предположительно, ещё до разделения клеточной жизни на три домена[56]. Это подтверждается тем, что некоторые вирусные белки не обнаруживают гомологии с белками бактерий, архей и эукариот, что свидетельствует о сравнительно давнем обособлении этой группы. В остальном же достоверно объяснить происхождение вирусов на основании трёх закрепившихся классических гипотез не удаётся, что делает необходимыми пересмотр и доработку этих гипотез[56].
Мир РНК[править | править код]
Гипотеза мира РНК[57] и компьютерный анализ последовательностей вирусной ДНК и ДНК хозяина дают лучшее понимание эволюционных взаимоотношений между различными группами вирусов и могут помочь определить предков современных вирусов. До настоящего времени такие исследования пока не прояснили, какая из трёх основных гипотез верна[57]. Однако представляется маловероятным, чтобы все современные вирусы имели общего предка, и, возможно, в прошлом вирусы независимо возникали несколько раз по одному или нескольким механизмам, так как между различными группами вирусов имеются значительные различия в организации генетического материала[58].

Препарат головного мозга коровы, поражённой коровьим бешенством. В сером веществе образуются микроскопические полости, которые придают ткани вид губки
Прионы[править | править код]
Прионы — это инфекционные белковые молекулы, не содержащие ДНК или РНК[59]. Они вызывают такие заболевания, как почесуха овец[60], губчатая энцефалопатия крупного рогатого скота и хроническая слабость (англ. chronic wasting disease) у оленей. К прионным болезням человека относят куру, болезнь Крейтцфельдта — Якоба и синдром Герстмана — Штраусслера — Шейнкера[61]. Прионы способны стимулировать образование собственных копий. Прионный белок способен существовать в двух изоформах: нормальной (PrPC) и прионной (PrPSc). Прионная форма, взаимодействуя с нормальным белком, способствует его превращению в прионную форму. Хотя прионы фундаментально отличаются от вирусов и вироидов, их открытие даёт больше оснований поверить в то, что вирусы могли произойти от самовоспроизводящихся молекул[62].
Биология[править | править код]
Вирусы как форма жизни[править | править код]
Пока вирус находится во внеклеточной среде или в процессе заражения клетки, он существует в виде независимой частицы. Вирусные частицы (вирионы) состоят из двух или трёх компонентов: генетического материала в виде ДНК или РНК (некоторые, например мимивирусы, имеют оба типа молекул); белковой оболочки (капсида), защищающей эти молекулы, и, в некоторых случаях, — дополнительных липидных оболочек. Наличие капсида отличает вирусы от вирусоподобных инфекционных нуклеиновых кислот — вироидов. В зависимости от того, каким типом нуклеиновой кислоты представлен генетический материал, выделяют ДНК-содержащие вирусы и РНК-содержащие вирусы; на этом принципе основана классификация вирусов по Балтимору. Ранее к вирусам также ошибочно относили прионы, однако впоследствии оказалось, что эти возбудители представляют собой особые инфекционные белки и не содержат нуклеиновых кислот. Форма вирусов варьирует от простой спиральной и икосаэдрической до более сложных структур. Размеры среднего вируса составляют около одной сотой размеров средней бактерии. Большинство вирусов слишком малы, чтобы быть отчётливо различимыми под световым микроскопом.
Вирусы являются облигатными паразитами, так как не способны размножаться вне клетки. Вне клетки вирусные частицы не проявляют признаки живого и ведут себя как частицы биополимеров. От живых паразитарных организмов вирусы отличаются полным отсутствием основного и энергетического обмена и отсутствием сложнейшего элемента живых систем — аппарата трансляции (синтеза белка), степень сложности которого превышает таковую самих вирусов.
Согласно одному из определений вирусы представляют собой форму жизни, согласно другому вирусы являются комплексами органических молекул, взаимодействующими с живыми организмами. Вирусы характеризуют как «организмы на границе живого»[21]. Вирусы похожи на живые организмы в том, что они имеют свой набор генов и эволюционируют путём естественного отбора[63], а также в том, что способны размножаться, создавая собственные копии путём самосборки. Вирусы имеют генетический материал, однако лишены клеточного строения, а именно эту черту обычно рассматривают как фундаментальное свойство живой материи. У вирусов нет собственного обмена веществ, и для синтеза собственных молекул им необходима клетка-хозяин. По этой причине они не способны размножаться вне клетки[64]. В то же время такие бактерии, как риккетсии и хламидии, несмотря на то, что не могут размножаться вне клеток хозяина, считаются живыми организмами[65][66]. Общепризнанные формы жизни размножаются делением клетки, в то время как вирусные частицы самопроизвольно собираются в инфицированной клетке. От роста кристаллов размножение вирусов отличается тем, что вирусы наследуют мутации и находятся под давлением естественного отбора. Самосборка вирусных частиц в клетке даёт дополнительное подтверждение гипотезы, что жизнь могла зародиться в виде самособирающихся органических молекул[2]. Опубликованные в 2013 году данные о том, что некоторые бактериофаги обладают собственной иммунной системой, способной к адаптации[67], являются дополнительным доводом в пользу определения вируса как формы жизни.
Структура[править | править код]

Строение икосаэдрических вирионов:
А. Вирус, не имеющий липидной оболочки (например, пикорнавирус).
B. Оболочечный вирус (например, герпесвирус).
Цифрами обозначены: (1) капсид, (2) геномная нуклеиновая кислота, (3) капсомер, (4) нуклеокапсид, (5) вирион, (6) липидная оболочка, (7) мембранные белки оболочки
Палочковидный вирион вируса табачной мозаики. Цифрами обозначены: (1) РНК-геном вируса, (2) капсомер, состоящий всего из одного протомера, (3) зрелый участок капсида
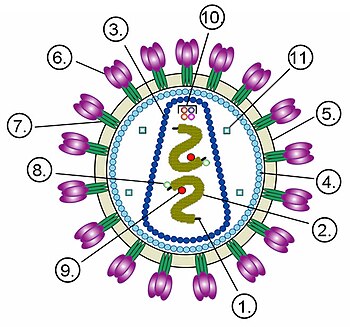
Строение вириона неикосаэдрического оболочечного вируса на примере ВИЧ. Цифрами обозначены: (1) РНК-геном вируса, (2) нуклеокапсид, (3) капсид, (4) белковый матрикс, подстилающий (5) липидную мембрану, (6) gp120 — гликопротеин, с помощью которого происходит связывание вируса с клеточной мембраной, (7) gp41 — гликопротеин.
Цифрами 8—11 обозначены белки, входящие в состав вириона и необходимые вирусу на ранних стадиях инфекции: (8) — интеграза, (9) — обратная транскриптаза, (10) — Vif, Vpr, Nef и p7, (11) — протеаза
Вирусы демонстрируют огромное разнообразие форм и размеров. Как правило, вирусы значительно мельче бактерий. Большинство изученных вирусов имеют диаметр в пределах от 20 до 300 нм. Некоторые филовирусы имеют длину до 1400 нм, но их диаметр составляет лишь 80 нм[68]. В 2013 году самым крупным из известных вирусов считался Pandoravirus размерами 1 × 0,5 мкм, однако в 2014 году из многолетней мерзлоты из Сибири был описан Pithovirus, достигающий 1,5 мкм в длину и 0,5 мкм в диаметре. В настоящий момент он считается крупнейшим из известных вирусов[69]. Большинство вирионов невозможно увидеть в световой микроскоп, поэтому используют электронные — как сканирующие, так и просвечивающие[70]. Чтобы вирусы резко выделялись на окружающем фоне, применяют электронно-плотные «красители». Они представляют собой растворы солей тяжёлых металлов, таких как вольфрам, которые рассеивают электроны на покрытой ими поверхности. Однако обработка такими веществами ухудшает видимость мелких деталей. В случае негативного контрастирования «окрашивается» только фон[71].
Зрелая вирусная частица, известная как вирион, состоит из нуклеиновой кислоты, покрытой защитной белковой оболочкой — капсидом. Капсид складывается из одинаковых белковых субъединиц, называемых капсомерами[72]. Вирусы могут также иметь липидную оболочку поверх капсида (суперкапсид), образованную из мембраны клетки-хозяина. Капсид состоит из белков, кодируемых вирусным геномом, а его форма лежит в основе классификации вирусов по морфологическому признаку[73][74]. Сложноорганизованные вирусы, кроме того, кодируют специальные белки, помогающие в сборке капсида. Комплексы белков и нуклеиновых кислот известны как нуклеопротеины, а комплекс белков вирусного капсида с вирусной нуклеиновой кислотой называется нуклеокапсидом. Форму капсида и вириона в целом можно механически (физически) исследовать при помощи сканирующего атомно-силового микроскопа[75][76].
Капсид[править | править код]
Классифицируют четыре морфологических типа капсидов вирусов: спиральный, икосаэдрический, продолговатый и комплексный.
Спиральный[править | править код]
Эти капсиды состоят из одного типа капсомеров, уложенных по спирали вокруг центральной оси. В центре этой структуры может находиться центральная полость или канал. Такая организация капсомеров приводит к формированию палочковидных и нитевидных вирионов: они могут быть короткими и очень плотными или длинными и очень гибкими. Генетический материал, как правило, представлен одноцепочечной РНК (в некоторых случаях одноцепочечной ДНК) и удерживается в белковой спирали ионными взаимодействиями между отрицательными зарядами на нуклеиновых кислотах и положительными зарядами на белках. В целом, длина спирального капсида зависит от длины окружённой им нуклеиновой кислоты, а диаметр определяется размером и расположением капсомеров. Примером спирального вируса может служить вирус табачной мозаики[77].
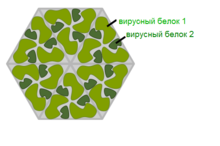
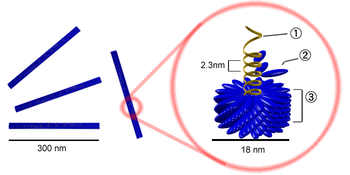
Вирус с капсомером типа гексон. Эта иллюстрация показывает, что вирусный капсид может быть выстроен из множественных копий всего лишь двух белков
Икосаэдрический[править | править код]
Большинство вирусов животных имеют икосаэдрическую или почти шарообразную форму с икосаэдрической симметрией. Правильный икосаэдр является оптимальной формой для закрытого капсида, сложенного из одинаковых субъединиц. Минимальное необходимое число одинаковых капсомеров — 12, каждый капсомер состоит из пяти идентичных субъединиц. Многие вирусы, такие как ротавирус, имеют более двенадцати капсомеров и выглядят круглыми, но сохраняют икосаэдрическую симметрию. Капсомеры, находящиеся в вершинах, окружены пятью другими капсомерами и называются пентонами. Капсомеры треугольных граней имеют 6 соседей-капсомеров и называются гексонами[78]. Гексоны, по существу, являются плоскими, а пентоны, образующие 12 вершин, — изогнутыми. Один и тот же белок может выступать субъединицей и пентомеров, и гексамеров, или же они могут состоять из различных белков.
Продолговатый[править | править код]
Продолговатыми называют икосаэдрические капсиды, вытянутые вдоль оси симметрии пятого порядка. Такая форма характерна для головок бактериофагов[79].
Комплексный[править | править код]
Форма этих капсидов ни чисто спиральная, ни чисто икосаэдрическая. Они могут нести дополнительные наружные структуры, такие как белковые хвосты или сложные наружные стенки. Некоторые бактериофаги, такие как фаг Т4, имеют комплексный капсид, состоящий из икосаэдрической головки, соединённой со спиральным хвостом, который может иметь шестигранное основание с отходящими от него хвостовыми белковыми нитями. Этот хвост действует наподобие молекулярного шприца, прикрепляясь к клетке-хозяину и затем впрыскивая в неё генетический материал вируса[80].
Оболочка[править | править код]
Некоторые вирусы окружают себя дополнительной оболочкой из модифицированной клеточной мембраны (плазматической или внутренней, такой как ядерная мембрана или мембрана эндоплазматического ретикулума). Этот дополнительный билипидный слой называется суперкапсидом, а шиповидные выступы на нём — пепломерами. Липидная оболочка вируса испещрена белками, кодируемыми вирусными геномом и геномом хозяина; сама же мембрана, а также любые её углеводные компоненты происходят целиком из клетки-хозяина. Таким образом формируют свою оболочку вирус гриппа и ВИЧ. Инфекционность большинства вирусов, имеющих оболочку, зависит именно от этой оболочки[81].
Поксвирусы представляют собой крупные сложноорганизованные вирусы с необычной морфологией. Генетический материал вируса связан с белками в центральной дисковидной структуре, известной как нуклеоид. Нуклеоид окружён мембраной и двумя боковыми телами неизвестной функции. Вирус имеет наружную оболочку с большим количеством белков на её поверхности. Весь вирион слегка плейоморфен (то есть способен изменять форму и размер в зависимости от условий) и может принимать форму от овальной до блоковидной[82]. Мимивирус является одним из крупнейших описанных вирусов и имеет икосаэдрический капсид диаметром 400—500 нм. Белковые филаменты, отходящие от поверхности вириона, достигают 100 нм в длину[83][84]. В 2011 году исследователи обнаружили ещё более крупный вирус на океаническом дне вблизи побережья Чили. Вирус, которому было дано временное название Megavirus chilensis, можно увидеть даже в обычный оптический микроскоп[85].
Геном[править | править код]
| Свойства | Параметры |
|---|---|
| Нуклеиновая кислота |
|
| Форма |
|
| Количество цепей |
|
| Полярность |
|

Реассортимент может стать причиной появления нового высокопатогенного штамма человеческого гриппа
Вирусы демонстрируют огромное количество вариантов организации генома; в этом смысле они более разнообразны, чем растения, животные, археи и бактерии. Существуют миллионы различных типов вирусов[7], но только примерно 5000 из них были детально описаны[6]. Генетический материал вируса может быть представлен либо ДНК, либо РНК, соответственно, вирусы подразделяют на ДНК-содержащие и РНК-содержащие. Подавляющее большинство вирусов являются РНК-содержащими. Вирусы растений чаще всего содержат одноцепочечную РНК, а бактериофаги, как правило, обладают двухцепочечными ДНК[86].
Вирусный геном может быть кольцевым, как у полиомавирусов, или линейным, как у аденовирусов. Форма генома не зависит от типа нуклеиновой кислоты. У многих РНК-содержащих вирусов и некоторых ДНК-содержащих вирусов геном часто представлен несколькими молекулами (частями), в связи с чем он называется сегментированным. У РНК-содержащих вирусов каждый сегмент часто кодирует только один белок, и обычно эти сегменты упаковываются в один капсид. Однако присутствие всех сегментов не всегда обязательно для инфекционности вируса, как это демонстрируют вирус мозаики костра[en] и некоторые другие вирусы растений[68].
Независимо от типа нуклеиновой кислоты вирусные геномы, как правило, относятся к одному из двух видов: либо одноцепочечному, либо двухцепочечному. Двухцепочечный геном включает пару комплементарных цепей нуклеиновой кислоты, а одноцепочечный — только одну цепь. И только у некоторых семейств (например, Hepadnaviridae) геном включает в себя как одноцепочечные, так и двухцепочечные участки[86].
Для большинства РНК-содержащих вирусов и некоторых вирусов с одноцепочечной ДНК определяют полярность нуклеиновой кислоты в зависимости от того, комплементарна ли она вирусной мРНК. Молекула РНК с положительной полярностью (плюс-цепь) имеет ту же последовательность нуклеотидов, что и мРНК, поэтому, по крайней мере, какая-то её часть может незамедлительно начать транслироваться клеткой-хозяином. РНК с отрицательной полярностью (минус-цепь) комплементарна мРНК, поэтому до начала трансляции на ней должна быть синтезирована положительная РНК при помощи фермента РНК-зависимой-РНК-полимеразы. Названия цепей ДНК для вирусов, содержащих одноцепочечную ДНК, сходны с таковыми для РНК: кодирующая цепь комплементарна мРНК (-), а некодирующая является её копией (+)[86]. Однако геномы нескольких типов ДНК- и РНК-содержащих вирусов представлены молекулами, имеющими различную полярность, то есть транскрипции может подвергаться любая цепь. Таковы, например, геминивирусы — вирусы растений, содержащие одноцепочечную ДНК, — и аренавирусы — вирусы животных с одноцепочечной РНК[87].
Размер генома широко варьирует у различных видов. Самым маленьким одноцепочечным ДНК-геномом обладает цирковирус из семейства Circoviridae: его геном кодирует лишь два белка и содержит всего 2000 нуклеотидов. Один из самых крупных геномов обнаружен у мимивируса: он содержит свыше 1,2 млн пар оснований и кодирует более тысячи белков[88]. Как правило, РНК-содержащие вирусы имеют меньший геном, чем ДНК-содержащие — размер их генома ограничен из-за большей вероятности ошибок во время репликации[40]. При большем размере генома ошибки, произошедшие во время его репликации, сделали бы вирус нежизнеспособным или неконкурентоспособным. Чтобы преодолеть это ограничение, РНК-вирусы часто имеют сегментированный геном — это уменьшает вероятность того, что ошибка в одном из сегментов окажется фатальной для всего генома. Напротив, ДНК-содержащие вирусы обычно имеют более крупные геномы благодаря большей точности их репликативных ферментов[89]. Однако вирусы, содержащие одноцепочечные ДНК, являются исключением из этого правила — скорость накопления мутаций в их геномах приближается к таковой для вирусов, содержащих одноцепочечные РНК[90].
Генетические изменения происходят у вирусов по разным механизмам. В их число входят случайные замены отдельных оснований в РНК или ДНК. В большинстве случаев эти точечные мутации являются «молчащими» — они не изменяют структуру белков, кодируемых мутантными генами, но иногда в результате таких изменений вирус может приобрести эволюционные преимущества, такие как устойчивость к противовирусным препаратам[91]. Дрейф антигенов имеет место тогда, когда в геноме вируса происходят масштабные изменения. Это может быть результатом рекомбинации или реассортимента. Когда это случается с вирусом гриппа, результатом может стать пандемия[92]. РНК-вирусы часто существуют как квазивиды или смесь вирусов одного вида, но с чуть различающимися нуклеотидными последовательностями генома. Такие квазивиды являются главной мишенью для естественного отбора[93].
Сегментированный геном даёт эволюционные преимущества: различные штаммы вируса с сегментированным геномом могут обмениваться генами и производить потомков с уникальными характеристиками. Это явление называется реассортиментом[94].
Генетическая рекомбинация — это процесс внесения разрыва в молекулу нуклеиновой кислоты с последующим «сшиванием» её с другими молекулами нуклеиновой кислоты. Рекомбинация может происходить между геномами двух вирусов, когда они заражают клетку одновременно. Исследования эволюции вирусов показали, что у изученных видов рекомбинация широко распространена[95]. Рекомбинация характерна как для РНК-, так и для ДНК-содержащих вирусов[96][97].
Жизненный цикл[править | править код]

Типичный жизненный цикл вируса на примере вируса гепатита C
Вирусы не размножаются клеточным делением, поскольку не имеют клеточного строения. Вместо этого они используют ресурсы клетки-хозяина для образования множественных копий самих себя, и их сборка происходит внутри клетки.
Условно жизненный цикл вируса можно разбить на несколько взаимоперекрывающихся этапов (обычно выделяют 6 этапов[98]):
- Прикрепление представляет собой образование специфичной связи между белками вирусного капсида и рецепторами на поверхности клетки-хозяина. Это специфичное связывание определяет круг хозяев вируса. Например, ВИЧ поражает только определённый тип человеческих лейкоцитов. Это связано с тем, что оболочечный гликопротеин вируса gp120 специфично связывается с молекулой CD4 — хемокиновым рецептором, который обычно встречается на поверхности CD4+ T-лимфоцитов. Этот механизм обеспечивает инфицирование вирусом только тех клеток, которые способны осуществить его репликацию. Связывание с рецептором может вызвать конформационные изменения белка оболочки (или белка капсида в случае безоболочечного вируса), что в свою очередь служит сигналом к слиянию вирусной и клеточной мембран и проникновению вируса в клетку.
- Проникновение в клетку. На следующем этапе вирусу необходимо доставить внутрь клетки свой генетический материал. Некоторые вирусы также переносят внутрь клетки собственные белки, необходимые для её реализации (особенно это характерно для вирусов, содержащих негативные РНК). Различные вирусы для проникновения в клетку используют разные стратегии: например, пикорнавирусы впрыскивают свою РНК через плазматическую мембрану, а вирионы ортомиксовирусов захватываются клеткой в ходе эндоцитоза и попадают в кислую среду лизосом, где происходит депротеинизация вирусной частицы, после чего РНК в комплексе с вирусными белками преодолевает лизосомальную мембрану и попадает в цитоплазму. Вирусы также различают по тому, где происходит их репликация: часть вирусов (например, те же пикорнавирусы) размножается в цитоплазме клетки, а часть (например, ортомиксовирусы) в её ядре. Процесс инфицирования вирусами клеток грибов и растений отличается от инфицирования клеток животных. Растения имеют прочную клеточную стенку, состоящую из целлюлозы, а грибы — из хитина, так что большинство вирусов могут проникнуть в них только после повреждения клеточной стенки[99]. Однако почти все вирусы растений (включая вирус табачной мозаики) могут перемещаться из клетки в клетку в форме одноцепочечных нуклеопротеиновых комплексов через плазмодесмы[100]. Бактерии, как и растения, имеют крепкую клеточную стенку, которую вирусу, чтобы попасть внутрь, приходится повредить. Но в связи с тем, что клеточная стенка бактерий намного тоньше таковой у растений, некоторые вирусы выработали механизм впрыскивания генома в бактериальную клетку через толщу клеточной стенки, при котором капсид остаётся снаружи[101].
- Лишение оболочек представляет собой процесс потери капсида. Это достигается при помощи вирусных ферментов или ферментов клетки-хозяина, а может быть и результатом простой диссоциации. В конечном счёте вирусная геномная нуклеиновая кислота освобождается.
- Репликация вирусов подразумевает, прежде всего, репликацию генома. Репликация вируса включает синтез мРНК ранних генов вируса (с исключениями для вирусов, содержащих положительную РНК), синтез вирусных белков, возможно, сборку сложных белков и репликацию вирусного генома, которая запускается после активации ранних или регуляторных генов. Вслед за этим может последовать (у комплексных вирусов с крупными геномами) ещё один или несколько кругов дополнительного синтеза мРНК: «поздняя» экспрессия генов приводит к синтезу структурных или вирионных белков.

Отпочковывание вируса от клетки-хозяина
- Вслед за этим происходит сборка вирусных частиц, позже происходят некоторые модификации белков. У вирусов, таких как ВИЧ, такая модификация (иногда называемая созреванием) происходит после выхода вируса из клетки-хозяина[102].
- Выход из клетки. Вирусы могут покинуть клетку после лизиса, процесса, в ходе которого клетка погибает из-за разрыва мембраны и клеточной стенки, если такая есть. Эта особенность есть у многих бактериальных и некоторых животных вирусов. Некоторые вирусы подвергаются лизогенному циклу, где вирусный геном включается путём генетической рекомбинации в специальное место хромосомы клетки-хозяйки. Тогда вирусный геном называется провирусом, или, в случае бактериофага, профагом[103]. Когда клетка делится, вирусный геном также удваивается. В пределах клетки вирус в основном не проявляет себя; однако в некоторый момент провирус или профаг может вызвать активацию вируса, который может вызвать лизис клеток-хозяев[104].
Активно размножающийся вирус не всегда убивает клетку-хозяина. Оболочечные вирусы, в том числе ВИЧ, обычно отделяются от клетки путём отпочковывания. В ходе этого процесса вирус обзаводится своей оболочкой, которая представляет собой модифицированный фрагмент клеточной мембраны хозяина или другой внутренней мембраны[105]. Таким образом, клетка может продолжать жить и продуцировать вирус.
Особенности жизненного цикла различных групп[править | править код]
Генетический материал внутри вирусных частиц и способ его репликации значительно отличается у различных вирусов.
- ДНК-содержащие вирусы. Репликация генома у большинства ДНК-содержащих вирусов происходит в клеточном ядре. Если клетка имеет соответствующий рецептор на своей поверхности, эти вирусы проникают в клетку либо путём непосредственного слияния с клеточной мембраной (напр. герпесвирусы), либо — что бывает чаще — путём рецептор-зависимого эндоцитоза. Большинство ДНК-содержащих вирусов полностью полагаются на синтетический аппарат клетки-хозяина для производства их ДНК и РНК, а также последующего процессинга РНК. Однако вирусы с крупными геномами (например, поксвирусы) могут сами кодировать большую часть необходимых для этого белков. Геному вируса эукариот необходимо преодолеть ядерную оболочку для того, чтобы получить доступ к ферментам, синтезирующим ДНК и РНК, в случае же бактерифагов ему достаточно просто проникнуть в клетку[106][107].
- РНК-содержащие вирусы. Репликация таких вирусов обычно происходит в цитоплазме. РНК-содержащие вирусы можно подразделить на 4 группы в зависимости от способа их репликации. Механизм репликации определяется тем, является ли геном вируса одноцепочечным или двухцепочечным, вторым важным фактором в случае одноцепочечного генома является его полярность (может ли он непосредственно служить матрицей для синтеза белка рибосомами). Все РНК-вирусы используют собственную РНК-репликазу для копирования своих геномов[108].
- Вирусы, использующие обратную транскрипцию. Эти вирусы содержат одноцепочечную РНК (Retroviridae, Metaviridae, Pseudoviridae) или двухцепочечную ДНК (Caulimoviridae и Hepadnaviridae). РНК-содержащие вирусы, способные к обратной транскрипции (ретровирусы, например, ВИЧ), используют ДНК-копию генома как промежуточную молекулу при репликации, а содержащие ДНК (параретровирусы, например, вирус гепатита B) — РНК[109]. В обоих случаях используется обратная транскриптаза, или РНК-зависимая ДНК-полимераза. Ретровирусы встраивают ДНК, образующуюся в процессе обратной транскрипции, в геном хозяина, такое состояние вируса называется провирусом. Параретровирусы же этого не делают, хотя встроенные копии их генома могут давать начало инфекционным вирусам, особенно у растений[110]. Вирусы, использующие обратную транскрипцию, восприимчивы к противовирусным препаратам, ингибирующим обратную транскриптазу, в том числе к зидовудину и ламивудину.
Действие на клетки[править | править код]

Диапазон структурных и биохимических эффектов, оказываемых вирусом на инфицированную клетку, очень широк[111]. Они называются цитопатическими эффектами[112]. Большинство вирусных инфекций приводят к гибели клеток-хозяев. Причинами гибели могут быть лизис клетки, изменения клеточной мембраны и апоптоз[113]. Часто причиной гибели клетки является подавление её нормальной активности белками вируса, не все из которых входят в состав вирусной частицы[114].
Некоторые вирусы не вызывают никаких видимых изменений в поражённой клетке. Клетки, в которых вирус находится в латентном состоянии и неактивен, имеют мало признаков инфекции и нормально функционируют[115]. Это является причиной хронических инфекций, и вирус при них может никак себя не проявлять многие месяцы или годы. Так часто бывает, например, с вирусом герпеса[116][117]. Некоторые вирусы, например вирус Эпштейна — Барр, могут вызывать быстрое размножение клеток без появления злокачественности[118], в то время как другие, такие как папилломавирусы, могут вызвать рак[119].
Диапазон хозяев[править | править код]
Вирусы, несомненно, являются самыми многочисленными биологическими объектами на Земле, и по этому показателю они превосходят все организмы, вместе взятые[120]. Они поражают все формы клеточных организмов, включая животных, растения, бактерии и грибы[6]. Тем не менее различные типы вирусов могут поражать только ограниченный круг хозяев, многие вирусы видоспецифичны. Некоторые, как, например, вирус оспы, могут поражать только один вид — людей[121], в подобных случаях говорят, что вирус имеет узкий диапазон хозяев. Напротив, вирус бешенства может поражать различные виды млекопитающих, то есть он имеет широкий диапазон хозяев[122]. Вирусы растений безвредны для животных, а большинство вирусов животных безопасны для человека[123]. Диапазон хозяев некоторых бактериофагов ограничивается одним штаммом бактерий, и они могут использоваться для определения штаммов, вызывающих вспышки инфекционных заболеваний методом фагового типирования[124].
Распространение[править | править код]
Вирусы распространяются многими способами: вирусы растений часто передаются от растения к растению насекомыми, питающимися растительными соками, к примеру, тлями; вирусы животных могут распространяться кровососущими насекомыми, такие организмы известны как переносчики. Вирус гриппа распространяется воздушно-капельным путём при кашле и чихании. Норовирус и ротавирус, обычно вызывающие вирусные гастроэнтериты, передаются фекально-оральным путём при контакте с заражённой пищей или водой. ВИЧ является одним из нескольких вирусов, передающихся половым путём и при переливании заражённой крови. Каждый вирус имеет определённую специфичность к хозяевам, определяющуюся типами клеток, которые он может инфицировать. Круг хозяев может быть узок или, если вирус поражает многие виды, широк[125].
Классификация[править | править код]
В таксономии живой природы вирусы выделяют в отдельный таксон, образующий в классификации Systema Naturae 2000 вместе с доменами Bacteria, Archaea и Eukaryota корневой таксон Biota[126]. В течение XX века в систематике выдвигались предложения о создании выделенного таксона для неклеточных форм жизни (Aphanobionta Novak, 1930[127]; надцарство Acytota Jeffrey, 1971[128]; Acellularia[129]), однако такие предложения не были кодифицированы[130].
Главной задачей классификации является описание разнообразия вирусов и группировка их на основании общих свойств. В 1962 году Андре Львов, Роберт Хорн[en] и Пауль Турнье были первыми, кто разработал основные принципы классификации вирусов на основании Линнеевской иерархической системы[131]. Основными таксонами в этой системе являются отдел, класс, порядок, семейство, род и вид. Вирусы были разделены на группы по их (а не их хозяев) общим свойствам и типу нуклеиновых кислоты в геноме[132]. Классификация вирусов по Балтимору является, по существу, дополнением к более традиционной классификации[133].
Систематику и таксономию вирусов в настоящий момент кодифицирует и поддерживает Международный комитет по таксономии вирусов (International Committee on Taxonomy of Viruses, ICTV), поддерживающий также и таксономическую базу (The Universal Virus Database, ICTVdB).
Развитие вирусной метагеномики привело к идентификации множества новых РНК-вирусов, которые помогли воссоздать эволюционную историю РНК-содержащих вирусов[133]. Не существует ни одного гена, который был бы общим для всех вирусов, что однозначно свидетельствует в пользу полифилетического происхождения вирусов. Тем не менее, существует набор из приблизительно 20 ключевых генов, кодирующих белки, которые участвуют в репликации вирусов и образовании вирионов. На основании сходства по этим генам в 2018 году было предложено разделение всех вирусов на таксоны высшего ранга — реалмы[комм. 2][134]. По состоянию на 2021 год вирусы подразделяются на шесть реалмов[134].
Классификация ICTV[править | править код]
Международный комитет по таксономии вирусов разработал современную классификацию вирусов и выделил основные свойства вирусов, имеющие больший вес для классификации с сохранением единообразия семейств.
Была разработана объединённая таксономия (универсальная система для классификации вирусов). Седьмой отчёт ICTV закрепил для первых пор понятие о виде вируса как о низшем таксоне в иерархии вирусов[135][комм. 4]. Однако к настоящему моменту была изучена лишь небольшая часть от общего разнообразия вирусов, анализ образцов вирусов из человеческого организма выявил, что около 20 % последовательностей вирусных нуклеиновых кислот ещё не было рассмотрено ранее, а образцы из окружающей среды, например, морской воды и океанского дна, показали, что подавляющее большинство последовательностей являются совершенно новыми[136].
Допустимыми таксономическими единицами являются следующие ранги[137], которым соответствуют определённые суффиксы в научных названиях таксонов[138]:
- Реалм (-viria) и субреалм (-vira)
- Царство (-virae) и подцарство (-virites)
- Тип (-viricota) и подтип (-viricotina)
- Класс (-viricetes) и подкласс (-viricetidae)
- Порядок (-virales) и подпорядок (-virineae)
- Семейство (-viridae) и подсемейство (-virinae)
- Род (-virus) и подрод (-virus)
- Вид (-virus)
- Род (-virus) и подрод (-virus)
- Семейство (-viridae) и подсемейство (-virinae)
- Порядок (-virales) и подпорядок (-virineae)
- Класс (-viricetes) и подкласс (-viricetidae)
- Тип (-viricota) и подтип (-viricotina)
- Царство (-virae) и подцарство (-virites)
Классификация ICTV не регулирует подвиды, штаммы и изоляты[139].
По состоянию на апрель 2021 года выделяют 6 реалмов, 10 царств, 17 типов, 2 подтипа, 39 классов, 59 порядков, 8 подпорядков, 189 семейств, 136 подсемейств, 2224 рода, 70 подродов и 9110 видов вирусов, вироидов и сателлитов[1]. Ещё свыше 3000 вирусов не классифицировано[140].
Как отмечалось ранее, реалмы были выделены на основании сравнений последовательностей приблизительно двадцати ключевых генов, которые кодируют белки, участвующие в репликации вирусов и образовании вирионов. Реалм Riboviria включает вирусы, кодирующие РНК-зависимую РНК-полимеразу; к нему относятся вирусы III, IV, V, VI и VII групп классификации по Балтимору. К реалму Riboviria относятся практически все вирусы, имеющие РНК-геном, за исключением вируса гепатита дельта и родственных ему вирусов, которые выделяются в реалм Ribozyviria. Вирусы реалма Ribozyviria похожи на вироиды и имеют рибозим, участвующий в созревании вирусной РНК, однако, в отличие от вироидов, они кодируют нуклеокапсидный белок. Вирусы, геномы которых представлены одноцепочечной ДНК, выделяют в реалм Monodnaviria. У большинства членов этого реалма имеется общий знаковый белок — эндонуклеаза, задействованная в репликации по типу катящегося кольца. Многие представители Monodnaviria также имеют капсидный белок, содержащий укладку jelly roll. Также к реалму Monodnaviria относят два семейства вирусов с геномами в виде двуцепочечной ДНК, Polyomaviridae и Papillomaviridae, которые произошли от вирусов с геномами в виде одноцепочечной ДНК, вероятно, вирусов семейства Parvoviridae. К реалму Varidnaviria относятся все вирусы с геномами в виде двуцепочечной ДНК, которые имеют основной капсидный белок (англ. major capsid protein) с укладкой jelly roll[141]. Реалм Duplodnaviria также включает вирусы с геномами в виде двуцепочечной ДНК, однако в их основных капсидных белках присутствует укладка HK97[142]. В реалм Adnaviria выделены вирусы, у которых геном в составе вириона представлен двуцепочечной ДНК в A-форме[134].
Классификация по Балтимору[править | править код]

Группы вирусов по Балтимору. Условные обозначения: оц — одноцепочечная, дц — двуцепочечная, РТ — ретровирус или ретроидный вирус
Лауреат Нобелевской премии биолог Дейвид Балтимор разработал классификацию вирусов, названную его именем[35][143]. Классификация ICTV в настоящее время объединяется с классификацией по Балтимору, составляя современную систему классификации вирусов[144][145].
Классификация вирусов по Балтимору основывается на механизме образования мРНК. Вирусы синтезируют мРНК из собственного генома для образования белков и репликации своей нуклеиновой кислоты, однако каждое семейство вирусов имеет собственный механизм этого процесса. Вирусные геномы могут быть одноцепоченые (оц) или двухцепочечные (дц), ДНК- или РНК-содержащие, могут использовать или не использовать обратную транскриптазу. Кроме того, одноцепочечные РНК-вирусы в составе своего генома могут иметь положительную (+) или отрицательную (-) цепь РНК.
Эта система включает в себя семь основных групп[143][146]:
- (I) Вирусы, содержащие двуцепочечную ДНК и не имеющие РНК-стадии (например, герпесвирусы, поксвирусы, паповавирусы, мимивирус).
- (II) Вирусы, содержащие одноцепочечную молекулу ДНК (например, парвовирусы). В этом случае ДНК всегда положительной полярности.
- (III) Вирусы, содержащие двуцепочечную РНК (например, ротавирусы).
- (IV) Вирусы, содержащие одноцепочечную молекулу РНК положительной полярности (например, пикорнавирусы, флавивирусы).
- (V) Вирусы, содержащие одноцепочечную молекулу РНК негативной или двойной полярности (например, ортомиксовирусы, филовирусы).
- (VI) Вирусы, содержащие одноцепочечную молекулу РНК положительной полярности и имеющие в своем жизненном цикле стадию синтеза ДНК на матрице РНК, ретровирусы (например, ВИЧ).
- (VII) Вирусы, содержащие частично двуцепочечную, частично одноцепочечную ДНК[147][148] и имеющие в своём жизненном цикле стадию синтеза ДНК на матрице РНК, ретроидные вирусы (например, вирус гепатита B)[149].
Дальнейшее деление производится на основе таких признаков, как структура генома (наличие сегментов, кольцевая или линейная молекула), генетическое сходство с другими вирусами, наличие липидной оболочки, таксономическая принадлежность организма-хозяина и других.
Эволюция[править | править код]
Не существует гена, который бы присутствовал у всех вирусов, поэтому вирусы в целом являются полифилетической группой. Развитие вирусной метагеномики привело к идентификации множества новых РНК-вирусов, которые помогли воссоздать эволюционную историю вирусов[133]. Тем не менее, существует набор из приблизительно 20 ключевых генов, кодирующих белки, которые участвуют в репликации вирусов и образовании вирионов. На основании сходства по этим генам в 2018 году было предложено разделение всех вирусов на таксоны высшего ранга — реалмы. Представители реалма Riboviria, который считается монофилетическим, могли произойти от древнего простого генетического элемента, который имел РНК-зависимую РНК-полимеразу. Вирусы, геномы которых представлены двуцепочечной РНК, скорее всего, произошли как минимум дважды от разных групп вирусов с одноцепочечной РНК положительной полярности. Группы IV (геном — одноцепочечная РНК положительной полярности) и V (геном — одноцепочечная РНК отрицательной полярности) в классификации по Балтимору являются монофилетическими, а группа III, к которой относятся вирусы с геномами в виде двуцепочечной РНК, — полифилетической. Однако вирус гепатита дельта и другие члены реалма Ribozyviria формально относятся к группе V, хотя фундаментально отличаются от других РНК-содержащих вирусов наличием рибозима, участвующего в репликации вирусной РНК, и потребности в вирусе-хозяине (вирусы-сателлиты), без которого их размножение невозможно. Если включать Ribozyviria в группу V, то её также следует считать полифилетической[134].
Вирусы, геномы которых представлены одноцепочечной ДНК, образуют группу II в классификации по Балтимору и реалм Monodnaviria. Несмотря на то что все они имеют эндонуклеазу, задействованную в репликации по типу катящегося кольца, эта группа вирусов является полифилетической и возникала в ходе эволюции несколько раз как объединение в одном генетическом элементе гена указанной эндонуклеазы, заимствованной у плазмид, и гена капсидного белка у различных вирусов IV группы в ходе рекомбинации. Эволюционный путь вирусов с геномом в виде двуцепочечной ДНК также непрост. Они подразделяются на три неродственных реалма: Duplodnaviria, Varidnaviria и Adnaviria. Вирусы с геномом в виде двуцепочечной ДНК в ходе эволюции возникали по меньшей мере четыре раза от независимых предков[134].
Роль в заболеваниях человека[править | править код]

Примерами наиболее известных вирусных заболеваний человека могут служить простуда (она может иметь и бактериальную этиологию), грипп, ветряная оспа и простой герпес. Многие серьёзные болезни, например, геморрагическая лихорадка Эбола, СПИД, птичий грипп и тяжёлый острый респираторный синдром тоже вызываются вирусами. Относительная способность вируса вызывать заболевание характеризуется термином вирулентность. Некоторые заболевания исследуются на наличие вирусов в числе вызывающих агентов, например, возможна связь между человеческим герпесвирусом 6 типа и нейрологическими заболеваниями, такими как рассеянный склероз и синдром хронической усталости[150], а борнавирус, возбудитель нейрологических заболеваний у лошадей, возможно, вызывает и психические расстройства у людей[151].
Вирусы различаются механизмами воздействия на организм хозяина, которые сильно зависят от вида. На клеточном уровне этот механизм предполагает лизис клетки, то есть её смерть. У многоклеточных организмов при гибели большого числа клеток начинает страдать организм в целом. Хотя вирусы подрывают нормальный гомеостаз, приводя к заболеванию, они могут существовать внутри организма и относительно безвредно. Некоторые вирусы (например, вирус простого герпеса первого типа) могут пребывать внутри тела человека в состоянии покоя, которое называется латентностью[152]. Оно характерно для вирусов герпеса, в том числе вируса Эпштейна — Барр, вызывающего инфекционный мононуклеоз, а также вируса, вызывающего ветрянку и опоясывающий лишай. Большинство людей переболело по крайней мере одним из этих типов вируса герпеса[153]. Однако такие латентные вирусы могут и принести пользу, поскольку присутствие этих вирусов может вызвать иммунный ответ против бактериальных патогенов, например, чумной палочки (Yersinia pestis)[154].
Некоторые вирусы могут вызывать пожизненные или хронические инфекции, при которых вирус продолжает размножаться в теле хозяина, несмотря на его защитные механизмы[155]. Так происходит, например, при инфекциях, вызванных вирусами гепатита B и C. Хронически больные люди (носители), таким образом, выступают в роли резервуара инфекции[156]. Если в популяции доля вирусоносителей высока, то такое состояние характеризуется как эпидемия[157].
Эпидемиология[править | править код]
Вирусная эпидемиология является частью медицинской науки, изучающей передачу и контроль вирусных инфекций среди людей. Передача вирусов может осуществляться вертикально, то есть от матери к ребёнку, или горизонтально, то есть от человека к человеку. Примерами вертикальной передачи могут служить вирус гепатита B и ВИЧ, при которых малыш рождается уже заражённым[158]. Другим, более редким, примером служит вирус ветрянки и опоясывающего лишая, который, хотя и вызывает относительно слабые инфекции среди взрослых людей, может оказаться смертельным для эмбрионов и новорождённых малышей[159].
Горизонтальная передача является наиболее распространённым механизмом распространения вируса в популяции. Передача может осуществляться: при передаче жидкостей организма при половом акте, например, у ВИЧ; через кровь при переливании заражённой крови или пользовании грязным шприцом, например, у вируса гепатита C; передаче слюны губами, например, у вируса Эпштейна — Барр; проглатывании заражённой воды или пищи, например, у норовируса; при вдыхании воздуха, в котором находятся вирионы, например, вирус гриппа; насекомыми, например, комарами, повреждающими кожу хозяина, например, лихорадка денге. Скорость передачи вирусной инфекции зависит от нескольких факторов, к которым относят плотность популяции, количество чувствительных людей (то есть не имеющих иммунитета)[160], качество здравоохранения и погоду[161].
Эпидемиология используется, чтобы приостановить распространение инфекции в популяции во время вспышки вирусного заболевания[162]. Предпринимаются контрольные меры, основанные на знании того, как распространяется вирус. Важно найти источник (или источники) вспышки и идентифицировать вирус. Когда вирус определён, бывает возможным остановить инфекцию при помощи вакцин. Если вакцины недоступны, могут быть эффективными санация и дезинфекция. Часто заражённых людей изолируют от остального общества, то есть вирус помещается в карантин[163]. Чтобы взять под контроль вспышки ящура в Великобритании в 2001 году, были зарезаны тысячи коров[164]. У большинства инфекций человека и животных есть инкубационный период, в течение которого не проявляется никаких симптомов инфекции[165]. Инкубационный период вирусных заболеваний может длиться от нескольких дней до недель[166]. Часто перекрывающийся с ним, но в основном следующий после инкубационного периода — период передачи инфекции, когда заражённый человек или животное является заразным и может заразить других людей или животных[166]. Этот период также известен для многих инфекций, и знание длины обоих периодов является важным для контролирования вспышек[167]. Если вспышка приводит к необычно высокому числу случаев заболевания в популяции или регионе, то она называется эпидемией. Если вспышки имеют широкое распространение, то говорят о пандемии[168].
Эпидемии и пандемии[править | править код]
Численность коренного населения Америки была сильно уменьшена заразными заболеваниями, в частности, оспой, завезёнными в Америку европейскими колонизаторами. По некоторым оценкам, иноземными болезнями после прибытия Колумба в Америку, было убито около 70 % от всего коренного населения. Урон, нанесённый этими болезнями аборигенам, помог европейцам вытеснить и покорить их[169].
Пандемия — это эпидемия всемирного масштаба. Эпидемия испанского гриппа 1918 года, продолжавшаяся до 1919 года, относится к 5 категории пандемий вируса гриппа. Она была вызвана чрезвычайно агрессивным и смертоносным вирусом гриппа A. Его жертвами часто становились здоровые взрослые люди, в отличие от большинства вспышек гриппа, которые поражали в основном детей и подростков, людей старшего поколения и других ослабленных людей[170]. По старым оценкам, испанский грипп унёс 40—50 млн жизней[171], а по современным оценкам эта цифра приближается к 100 млн, то есть 5 % населения Земли в то время[172].
Большинство исследователей полагают, что ВИЧ появился в Африке южнее Сахары в течение XX столетия[173]. Сейчас эпидемия СПИД имеет масштаб пандемии. По оценкам, сейчас 38,6 миллионов людей на земле заражено ВИЧ[174]. По оценкам Объединённой программы Организации Объединённых Наций по ВИЧ/СПИД и Всемирной Организации Здравоохранения, от СПИДа (последней стадии ВИЧ-инфекции) умерло более 25 миллионов человек с момента регистрации первого случая заболевания 5 июня 1981 года, что делает его одной из наиболее разрушительных эпидемий за всю документированную историю[175]. В 2007 году было зарегистрировано 2,7 млн случаев заражения ВИЧ и 2 млн смертей от связанных с ВИЧ заболеваний[176].
Несколько высоколетальных вирусных патогенов относятся к семейству филовирусов (Filoviridae). Филовирусы представляют собой филаментовидные вирусы, которые вызывают геморрагическую лихорадку, к ним также относят возбудителя геморрагической лихорадки Эбола и марбургский вирус. Марбургский вирус привлёк широкое внимание прессы в апреле 2005 года из-за вспышки в Анголе. Продолжавшаяся с октября 2004 года и вплоть до 2005 года, эта вспышка вошла в историю как наиболее ужасная эпидемия любой геморрагической лихорадки[177].

Злокачественные опухоли[править | править код]
Вирусы могут вызывать злокачественные новообразования (в частности, гепатоцеллюлярную карциному или саркому Капоши) у человека и других видов, хотя они возникают лишь у небольшой части инфицированных. Опухолеродные вирусы относятся к различным семействам; они включают и РНК-, и ДНК-содержащие вирусы, поэтому единого типа «онковирус» не существует (устаревший термин, первоначально применявшийся для быстро трансформирующихся ретровирусов). Развитие рака определяется множеством факторов, такими как иммунитет хозяина[178] и его мутации[179]. К вирусам, способным вызывать рак у человека, относят некоторых представителей папилломавируса человека, вируса гепатита B и C, вируса Эпштейна — Барр, герпесвируса саркомы Капоши и человеческого T-лимфотропного вируса. Совсем недавно открытым вирусом рака человека является полиомавирус (полиомавирус клеток Меркеля), который в большинстве случаев вызывает редкую форму рака кожи, называемого карциномой клеток Меркеля[180]. Вирусы гепатита могут вызвать хроническую вирусную инфекцию, которая приводит к раку печени[181][182]. Заражение человеческим T-лимфотрофным вирусом может привести к тропическому спастическому параперезу и зрелой лейкемии Т-клеток[183]. Человеческие папилломавирусы могут вызывать рак шейки матки, кожи, ануса и полового члена[184]. Из герпесвирусов герпесвирус саркомы Капоши вызывает саркому Капоши и лимфому полости тела, вирус Эпштейна — Барр — лимфому Беркитта, лимфогранулематоз, нарушения B-лимфопролиферации и назофарингеальную карциному[185]. Полиомавирус клеток Меркеля близок к вирусу SV40 и полиомавирусам мышей, которые более 50 лет использовались как животные модели для изучения вирусного рака[186].
Защитная реакция хозяина[править | править код]
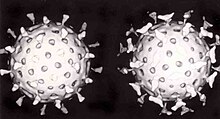
Первой защитной линией организма против вируса является врождённый иммунитет. Он включает клетки и другие механизмы, обеспечивающие неспецифическую защиту. Это значит, что клетки врождённого иммунитета распознают и реагируют на патогены общими способами, одинаково по отношению ко всем патогенам, но, в отличие от приобретённого иммунитета, врождённый иммунитет не даёт продолжительной и надёжной защиты хозяину[187].
Важным врождённым способом защиты организма эукариот против вирусов является РНК-интерференция[188]. Стратегия репликации многих вирусов предполагает наличие стадии двуцепочечной РНК. Для борьбы с подобными вирусами, клетка имеет систему неспецифической деградации одно- и двуцепочечных РНК. Когда подобный вирус попадает в клетку и освобождает геномную РНК в цитоплазму, белковый комплекс Dicer связывает и разрывает вирусную РНК на короткие фрагменты. Активируется биохимический путь, называемый RISC, который разрушает вирусную РНК, и препятствует размножению вируса. Ротавирусам удаётся избежать РНК-интерференции, сохраняя часть капсида даже внутри клетки и выпуская новообразованные мРНК через поры во внутреннем капсиде. Геномная двуцепочечная РНК же остаётся внутри него[189][190].
Когда система приобретённого иммунитета у позвоночных сталкивается с вирусом, она образует специфические антитела, присоединяющиеся к вирусу и часто делающие его неопасным. Это называется гуморальным иммунитетом. Наиболее важными являются два типа антител. Первый, называемый IgM, обладает высокой эффективностью в нейтрализации вирусов, но образуется клетками иммунной системы лишь в течение нескольких недель. Синтез второго — IgG — продолжается неопределённо долго. Присутствие IgM в крови хозяина говорит о наличии острой инфекции, в то время IgG свидетельствует об инфекции, перенесённой в прошлом[191]. Именно количество IgG измеряется при тестах на иммунитет[192]. Антитела могут продолжать оставаться эффективным защитным механизмом даже тогда, когда вирусу удаётся проникнуть в клетку. Клеточный белок TRIM21 может прикреплять антитела к поверхности вирусных частиц. Это вызывает последующее разрушение вирусной частицы ферментами клеточной протеосомной системы[193].
Второй защитный механизм позвоночных против вирусов называется клеточным иммунитетом и включает иммунные клетки, известные как T-лимфоциты. Клетки тела постоянно несут короткие фрагменты собственных белков на своих поверхностях, и, если Т-лимфоциты распознают здесь подозрительные вирусные фрагменты, клетка-хозяин разрушается клетками, называемыми Т-киллерами, и начинается образование специфичных к вирусу T-лимфоцитов. Такие клетки, как макрофаги, специализируются на презентации антигенов[194]. Важной защитной реакцией хозяина является продукция интерферона. Интерферон — это гормон, образуемый организмом в ответ на присутствие вируса. Его роль в иммунитете — комплексная, в конце концов он останавливает вирус, прекращая образование новых вирусов поражёнными клетками, убивая их и их близких соседей[195].
Не против всех вирусов образуется такой защитный иммунный ответ. ВИЧ удаётся избежать иммунного ответа, постоянно меняя последовательность аминокислот поверхностных белков вириона. Такие устойчивые вирусы уходят от иммунной системы, изолируясь от иммунных клеток, блокируя презентацию антигенов, благодаря устойчивости к цитокинам, уклоняясь от естественных киллеров, останавливая апоптоз клеток-хозяев, а также за счёт антигенной изменчивости[196]. Другие вирусы, называемые нейротропными вирусами, распространяются среди нервных клеток, то есть там, где иммунная система не в состоянии добраться до них из-за ГЭБ.
Профилактика и лечение[править | править код]
Так как вирусы используют для размножения естественные метаболические пути клеток-хозяев, их сложно уничтожить без применения препаратов, токсичных для самих клеток-хозяев. Наиболее эффективными медицинскими мерами против вирусных инфекций являются вакцинации, создающие иммунитет к инфекции, и противовирусные препараты, избирательно ингибирующие репликацию вирусов.
Вакцины[править | править код]
Вакцинация представляет собой дешёвый и эффективный способ предотвращения вирусных инфекций. Вакцины для предотвращения вирусных инфекций применялись ещё задолго до открытия самих вирусов. Их применение вызвано тяжёлым перенесением и смертностью от вирусных инфекций, таких как полиомиелит, корь, свинка и краснуха, поэтому лучше сделать прививку, чем переболеть[197]. С помощью вакцинации оспа была искоренена[198]. С помощью вакцин можно предотвратить более 30 вирусных инфекций у человека[199], а ещё больше вакцин используется для предотвращения вирусных заболеваний животных[200]. Вакцины могут включать ослабленные и убитые вирусы, а также вирусные белки (антигены)[201]. Живые вакцины содержат ослабленные формы вирусов, которые не вызывают болезни, но тем не менее вызывают иммунный ответ. Такие вирусы называются аттенуированными. Живые вакцины могут быть опасны для людей со слабым иммунитетом (то есть имеющих иммунодефицит), так как даже ослабленный вирус у них может вызвать исходное заболевание[202]. Для производства т. н. субъединичных вакцин используются биотехнологии и генная инженерия. В таких вакцинах используются только капсидные белки вирусов. Примером такой вакцины может служить вакцина против вируса гепатита B[203]. Субъединичные вакцины безвредны для людей с иммунодефицитом, так как они не могут вызвать заболевание[204]. Вакцина против вируса жёлтой лихорадки, содержащая ослабленный штамм 17D, пожалуй, является наиболее эффективной и безопасной из когда-либо созданных вакцин[205].
Противовирусные препараты[править | править код]


Противовирусные препараты часто представляют собой аналоги нуклеозидов. Они встраиваются в геном вируса в ходе репликации, и на этом жизненный цикл вируса останавливается, поскольку новосинтезированная ДНК неактивна. Это вызвано тем, что у аналогов отсутствуют гидроксильные группы, которые вместе с атомами фосфора соединяются и формируют жёсткий «остов» молекулы ДНК. Это называется цепной терминацией ДНК[206]. Примеры аналогов нуклеозидов — ацикловир, применяющийся против инфекций, вызванных простым вирусом герпеса, и ламивудин (против ВИЧ и вируса гепатита B). Ацикловир — один из старейших и наиболее часто назначаемых противовирусных препаратов[207]. Другие используемые противовирусные препараты имеют мишенью различные стадии жизненного цикла вирусов. Вирусу иммунодефицита человека, чтобы стать полностью заразным, необходим протеолитический фермент, известный как ВИЧ-1 протеаза. На основании этого разработан большой класс препаратов, называемых протеазными ингибиторами, инактивирующими этот фермент.
Гепатит C вызывается РНК-содержащим вирусом. У 80 % заражённых людей инфекция имеет хронический характер, и без лечения они останутся заражёнными до конца своих дней. Однако сейчас используется эффективное лекарство, состоящее из нуклеозидного аналога рибавирина, комбинированного с интерфероном[208]. Для лечения хронических носителей гепатита B было разработано схожее лечение с использованием ламивудина[209].
Вирусные заболевания у различных организмов[править | править код]
Вирусы поражают всю клеточную жизнь, но, несмотря на повсеместное распространение вирусов, каждый вид клеточных организмов имеет свой ряд поражающих вирусов, часто поражающих только этот вид[210]. Некоторые вирусы, называемые сателлитами, могут размножаться только в клетках, уже заражённых другим вирусом[53].
Вирусы животных[править | править код]
У животных вирусные инфекции вызывают иммунный ответ, который чаще всего приводит к уничтожению болезнетворного вируса. Иммунный ответ также можно вызвать вакцинами, дающими активный приобретённый иммунитет против конкретной вирусной инфекции. Однако некоторым вирусам, в том числе вирусу иммунодефицита человека и возбудителям вирусных гепатитов, удаётся ускользнуть от иммунного ответа, вызывая хроническую болезнь. Антибиотики не действуют на вирусы, однако было разработано несколько противовирусных препаратов (см. выше).
Вирусы являются важными патогенами домашнего скота. Вирусами вызываются такие заболевания, как ящур и «синий язык» (англ. bluetongue)[211]. Домашние животные, например, кошки, собаки и лошади, если их не вакцинировать, являются чувствительными к серьёзным вирусным болезням. Собачий парвовирус — это маленький ДНК-содержащий вирус, часто оказывается смертельным для щенков[212]. Однако большинство вирусов безвредно сосуществуют со своими хозяевами, не подавая никаких признаков или симптомов болезни[5].
Вирусы беспозвоночных[править | править код]
На долю беспозвоночных приходится около 80 % всех известных видов животных, поэтому нет ничего удивительного в том, что они скрывают в себе огромное множество вирусов различных типов. Наиболее изучены вирусы, поражающие насекомых, но даже здесь доступная по ним информация носит фрагментарный характер. Впрочем, в последнее время были описаны вирусные заболевания и у других беспозвоночных. Эти вирусы остаются малоизученными, и некоторые сообщения об открытии следует принимать с осторожностью, пока вирусная природа этих болезней не будет окончательно доказана. Кроме того, необходимо также проверить инфективность изолированных вирусов по отношению к неинфицированным хозяевам того же вида, у кого эти вирусы были обнаружены[213].
В настоящее время выделено отдельное семейство вирусов, поражающих главным образом членистоногих, в особенности — насекомых, обитающих в водных и влажных средах: иридовирусы (Iridoviridae, от англ. Invertebrate iridescent viruses — «радужные вирусы беспозвоночных»; такой цвет наблюдается у образцов поражённых насекомых). Они представляют собой икосаэдрические частицы 120—180 нм в диаметре, содержащие внутреннюю липидную мембрану и геном в виде двуцепочечной ДНК, содержащей 130—210 тыс. пар нуклеотидов[214].
Другие вирусы, поражающие насекомых: семейство Baculoviridae, подсемейство Entomopoxvirinae семейства Poxviridae, род Densovirus семейства Parvoviridae, некоторые вирусы семейств Rhabdoviridae, Reoviridae, Picornaviridae[215].
Как и все беспозвоночные, медоносная пчела чувствительна ко многим вирусным инфекциям[216].
Вирусы растений[править | править код]

Перцы, поражённые вирусом пятнистости
Существует много типов вирусов растений. Часто они вызывают снижение урожайности, принося большие убытки сельскому хозяйству, поэтому контроль таких вирусов очень важен с экономической точки зрения[217]. Вирусы растений часто распространяются от растения к растению организмами, известными как переносчики. Обычно ими выступают насекомые, но ими могут быть также грибы, черви-нематоды и одноклеточные организмы. Если контроль вируса растений признаётся экономически выгодным, например, в случае многолетних фруктовых деревьев, усилия направляются на устранение переносчиков или альтернативных хозяев, к примеру, сорняков[218]. Вирусы растений не могут поражать человека и других животных, так как они могут размножаться лишь в живых растительных клетках[219].
Растения имеют сложные и эффективные механизмы защиты от вирусов. Наиболее эффективным механизмом является наличие так называемого гена устойчивости (R от англ. resistance — «устойчивость»). Каждый R-ген отвечает за устойчивость к отдельному вирусу и вызывает гибель клеток, соседних с поражённой, что невооружённым глазом видно как большое пятно. Это останавливает развитие болезни вследствие остановки распространения вируса[220]. Другим эффективным методом является РНК-интерференция[221]. Будучи поражёнными вирусом, растения часто начинают вырабатывать природные противовирусные вещества, такие как салициловая кислота, оксид азота NO и активные формы кислорода[222].
Вирусы растений и созданные на их основе вирусоподобные частицы (VLPs) нашли применение в биотехнологиях и нанотехнологиях. Капсиды большинства вирусов растений имеют простую и устойчивую структуру, и вирусные частицы могут производиться в огромных количествах как поражённым растением, так и различными гетерологичными системами. Вирусы растений могут изменяться химически и генетически, заключая в оболочку инородные частицы, а также способны встраиваться в надмолекулярные структуры, что делает возможным их применение в биотехнологиях[223].
Для повышения достоверности результатов диагностики вирусологического статуса растений необходимо использовать как минимум два метода, причем, желательно, высокочувствительные — ИФА и ПЦР. Выявляемость вирусов повышается за счет использования гидроксипроизводного бензойной кислоты (ГПБК) в качестве эффективного антиоксиданта, учёта биологических особенностей культур и условий окружающей среды[224].
Вирусы грибов[править | править код]
Вирусы грибов называются миковирусами. В настоящий момент вирусы выделены у 73 видов из 57 родов, относящихся к 5 классам[225], но, предположительно, в безвредном состоянии вирусы существуют у большинства грибов. В общем эти вирусы представляют собой круглые частицы 30—45 нм диаметром, состоящие из множества субъединиц единственного белка, сложенных вокруг генома, представленного двуцепочечной РНК. Как правило, вирусы грибов относительно безвредны. Некоторые грибные штаммы могут поражаться многими вирусами, но большинство миковирусов тесно связаны со своим единственным хозяином, от которого передаются его потомкам. Классификацией вирусов грибов сейчас занимается специально созданный комитет в составе ICTV[225]. В данный момент он признаёт 3 семейства вирусов грибов, а наиболее изученные миковирусы относятся к семейству Totiviridae[226]
Установлено, что антивирусная активность пенициллиновых грибов вызвана индукцией интерферона двухцепочечной РНК от вирусов, поражающих грибы[225].
Если же вирус, попадая в гриб, проявляет свою вирулентность, то реакция гриба на это может быть различной: снижение или повышение вирулентности у патогенных видов, дегенерация мицелия и плодовых тел, изменение окраски, подавление спороношения. Некапсидированные вирусные РНК передаются через анастомозы независимо от митохондрий.
Вирусные заболевания могут наносить ущерб грибоводческим предприятиям, например, вызывать побурение плодовых тел шампиньона, изменение окраски у зимнего опёнка, что снижает их коммерческую ценность. Вирусы, вызывающие гиповирулентность грибов-патогенов, могут использоваться для борьбы с заболеваниями растений[227][228].
Вирусы протистов[править | править код]
К вирусам протистов относят вирусы, поражающие одноклеточных эукариот, не включённых в царство животные, растения или грибы. Некоторые из известных на данный момент вирусов протистов[229]:
| Название вируса (род) | Систематическое положение (семейство) |
Поражаемый протист |
|---|---|---|
| Dinornavirus | Alvernaviridae | Heterocapsa circularisquama |
| Endornavirus | Endornaviridae | Phytophthora |
| Labyrnavirus | Labyrnaviridae | Aurantiochytrium |
| Marnavirus | Marnaviridae | Heterosigma akashiwo |
| Marseillevirus | Marseilleviridae | Amoeba |
| Mimivirus | Mimiviridae | Acanthamoeba polyphaga |
| Chlorovirus | Phycodnaviridae | Paramecium bursaria |
| Coccolithovirus | Phycodnaviridae | Emiliania huxleyi |
| Prasinovirus | Phycodnaviridae | Micromonas pusilla |
| Prymnesiovirus | Phycodnaviridae | Chrysochromulina brevifilum |
| Raphidovirus | Phycodnaviridae | Heterosigma akashiwo |
| Cryspovirus | Partitiviridae | Cryptosporidium parvum |
| Hemivirus | Pseudoviridae | Volvox carteri |
| Pseudovirus | Pseudoviridae | Physarum polycephalum |
| Mimoreovirus | Reoviridae | Micromonas pusilla |
| Giardiavirus | Totiviridae | Giardia lamblia |
| Leishmaniavirus | Totiviridae | Leishmania |
| Trichomonasvirus | Totiviridae | Trichomonas vaginalis |
| Bacilladnavirus | Не определено | Chaetoceros salsugineum
Rhizosolenia setigera |
| Dinodnavirus | Не определено | Heterocapsa circularisquama |
| Rhizidiovirus | Не определено | Rhizidiomyces |
Многие вирусы простейших имеют необычно большие размеры. Например, геном Marseillevirus, впервые выделенный из амёбы, имеет геном размером 368 КБ, а Mamavirus, поражающий протиста Acanthamoeba, по размеру превосходит даже мимивирус (а его капсид достигает около 500 нм в диаметре) и некоторые бактерии. Также в число гигантских вирусов входит вирус, поражающий широко распространённого морского протиста Cafeteria roenbergensis (англ. Cafeteria roenbergensis virus, CroV)[230].
Вирусы бактерий[править | править код]
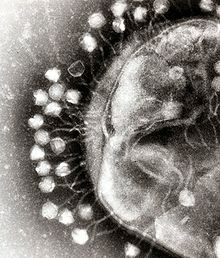
Бактериофаги представляют собой широко распространённую и разнообразную группу вирусов, достигающую большей численности в водных средах обитания — в океанах этих вирусов более чем в 10 раз больше, чем бактерий[231], достигая численности в 250 млн. вирусов на миллилитр морской воды[232]. Эти вирусы поражают специфичные для каждой группы бактерии, связываясь с клеточными рецепторами на поверхности клетки и затем проникая внутрь неё. В течение короткого промежутка времени (иногда считанных минут) бактериальная полимераза начинает транслировать вирусную мРНК в белки. Эти белки или входят в состав вирионов, собираемых внутри клетки, или являются вспомогательными белками, помогающими сборке новых вирионов, или вызывают лизис клетки. Вирусные ферменты вызывают разрушение клеточной мембраны, и, в случае фага Т4, всего лишь через 20 минут после проникновения в клетку на свет появляются свыше трёх сотен бактериофагов[233].
Главным механизмом защиты бактериальных клеток от бактериофагов является образование ферментов, разрушающих чужеродную ДНК. Эти ферменты, называемые эндонуклеазами рестрикции, «разрезают» вирусную ДНК, впрыснутую внутрь клетки[234]. Бактерии также используют систему, называемую CRISPR, которая хранит информацию о геномах вирусов, с которыми бактерия сталкивалась ранее, и это позволяет клетке блокировать репликацию вируса с помощью интерференции РНК[235][236]. Эта система обеспечивает приобретённый иммунитет бактериальной клетки.
Бактериофаги могут выполнять и полезную для бактерий функцию, так, именно бактериофаг, заражающий дифтерийные палочки, кодирует ген их токсина, нужного этим бактериям и столь опасного для человека[237]:45.
Вирусы архей[править | править код]
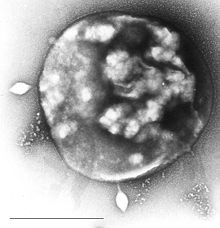
Некоторые вирусы размножаются внутри архей: это двуцепочечные ДНК-содержащие вирусы с необычной, подчас уникальной формой[8][243]. Наиболее детально они изучены у термофильных архей, в частности, порядков Sulfolobales и Thermoproteales[244]. Мерами защиты против этих вирусов могут быть РНК-интерференция от повторяющихся последовательностей ДНК в геномах архей, родственных генам вирусов[245][246].
Вирусы вирусов[править | править код]

При изучении вирусных фабрик мимивируса было обнаружено, что на них собираются небольшие вирионы и другого вируса, который был назван Спутником[247]. Спутник, по всей видимости, сам не способен заражать клетки амёб (которые служат хозяевами мимивируса) и размножаться в них, но может делать это совместно с мама- или мимивирусом, что классифицирует его как вирус-сателлит. Спутник стал первым известным вирусом-сателлитом, содержащим двухцепочечную ДНК и размножающимся в эукариотических клетках. Однако авторы работы предлагают рассматривать его не просто как сателлит, а как вирофаг (вирус вируса) по аналогии с бактериофагами (вирусами бактерий)[248][249][250]. Репликация как вирусов-сателлитов, так и вирофагов зависит от другого вируса и клетки-хозяина. Однако для репликативного цикла вирофагов характерны три уникальные особенности. 1) Отсутствует ядерная фаза репликации. 2) Репликация вирофагов происходит в вирусных фабриках гигантских ДНК-содержащих вирусов-хозяев. 3) Вирофаги зависят от ферментов, синтезируемых вирусами-хозяевами, но не клетками-хозяевами. Таким образом, вирофаги считаются паразитами гигантских ДНК-содержащих вирусов, например, мимивирусов и фикоднавирусов[251][252]. При этом синтез капсидных белков вирофагов (как и синтез белков всех известных вирусов) полностью зависит от трансляционного аппарата клетки-хозяина[253]. Хотя строгого доказательства ещё нет, некоторые факты говорят в пользу того, что Спутник действительно является вирофагом. Например, в его геноме присутствуют регуляторные элементы, характерные для мимивируса и узнаваемые его транскрипционным аппаратом (последовательности, близкие к позднему промотору мимивируса, сигналы полиаденилирования). Кроме того, присутствие Спутника снижает продуктивность размножения мимивируса: лизис клетки-хозяина происходит с задержкой, и образуются дефектные вирионы мимивируса[247].
По данным на 2016 год, из культивируемых клеток было изолировано пять вирофагов. Ещё 18 вирофагов описано на основе данных метагеномного анализа (геномы двух из них почти полностью секвенированы)[254][255].
Роль вирусов в биосфере[править | править код]
Вирусы являются самой распространённой формой существования органической материи на планете по численности. Они играют важную роль в регуляции численности популяций некоторых видов живых организмов (например, вирус дикования с периодом в несколько лет сокращает численность песцов в несколько раз).
Иногда вирусы образуют с животными симбиоз[256][257]. Так, например, яд некоторых паразитических ос содержит структуры, называемые поли-ДНК-вирусами (Polydnavirus, PDV), имеющие вирусное происхождение.
Однако основная роль вирусов в биосфере связана с их деятельностью в водах океанов и морей.
Роль в водных экосистемах[править | править код]
Вирусы — это самая распространенная форма жизни в океане, их концентрация достигает 10 млн вирусов на 1 миллилитр поверхности моря[258]. Чайная ложка морской воды содержит около миллиона вирусов[259]. Они необходимы для регуляции пресноводных и морских экосистем[260]. Большая часть этих вирусов является бактериофагами, безвредными для растений и животных. Они поражают и разрушают бактерии в водном микробном сообществе, таким образом, участвуя в важном процессе круговорота углерода в морской среде. Органические молекулы, освободившиеся из бактериальных клеток благодаря вирусам, стимулируют рост новых бактерий и водорослей[261].
Микроорганизмы составляют более 90 % биомассы в море. По оценкам, каждый день вирусы убивают около 20 % этой биомассы, а количество вирусов в океанах в 15 раз превышает число бактерий и архей. Вирусы являются главными агентами, вызывающими быстрое прекращение цветения воды[262], убивающего другую жизнь в море[263], за счёт гибели вызывающих его водорослей. Численность вирусов убывает с удалением от берега и с увеличением глубины, поскольку там меньше организмов-хозяев[264].
Значение морских вирусов очень велико. Регулируя процесс фотосинтеза, они играют второстепенную роль в сокращении количества углекислого газа в атмосфере приблизительно на 3 гигатонны углерода в год[264].
Как и другие организмы, морские млекопитающие восприимчивы к вирусным инфекциям. В 1988 и 2002 годах тысячи обыкновенных тюленей были убиты парамиксовирусом Phocine distemper virus[265]. В популяциях морских млекопитающих циркулирует множество других вирусов, в том числе калицивирусы, герпесвирусы, аденовирусы и парвовирусы[264].
Роль в эволюции[править | править код]
Вирусы являются важным естественным средством переноса генов между различными видами, что вызывает генетическое разнообразие и направляет эволюцию[38]. Считается, что вирусы сыграли центральную роль в ранней эволюции, ещё до расхождения бактерий, архей и эукариот, во времена последнего универсального общего предка жизни на Земле[266]. Вирусы и по сей день остаются одним из крупнейших живых хранилищ неисследованного генетического разнообразия на Земле[264].
Вирусы имеют генетические связи с представителями флоры и фауны Земли. Согласно последним исследованиям[источник не указан 2591 день], геном человека более чем на 32 % состоит из вирусоподобных элементов, транспозонов и их остатков. С помощью вирусов может происходить так называемый горизонтальный перенос генов (ксенология), то есть передача генетической информации не от непосредственных родителей к своему потомству, а между двумя неродственными (или даже относящимися к разным видам) особями. Так, в геноме высших приматов существует ген, кодирующий белок синцитин, который, как считается, был привнесён ретровирусом.
Применение[править | править код]
В науках о жизни и медицине[править | править код]

Вирусы имеют важное значение для исследований в молекулярной и клеточной биологии, так как они представляют собой простые системы, которые можно использовать для управления и изучения функционирования клеток[267]. Изучение и использование вирусов дало ценную информацию о различных аспектах клеточной биологии[268]. К примеру, вирусы применялись в генетических исследованиях, и они помогли нам прийти к пониманию ключевых механизмов молекулярной генетики, таких как репликация ДНК, транскрипция, процессинг РНК, трансляция, транспорт белков.
Генетики часто используют вирусы как векторы для ввода генов в изучаемые клетки. Это позволяет заставить клетку производить чуждые вещества, а также изучить эффект от ввода нового гена в геном. Аналогично в виротерапии вирусы используют как векторы для лечения различных болезней, так как они избирательно действуют на клетки и ДНК. Это даёт надежды, что вирусы смогут помочь в борьбе с раком и найдут своё применение в генотерапии. Некоторое время восточноевропейские учёные прибегали к фаговой терапии как к альтернативе антибиотикам, и интерес к таким методам возрастает, поскольку в настоящее время у некоторых патогенных бактерий обнаружена высокая устойчивость к антибиотикам[269].
Биосинтез заражёнными клетками чужеродных белков лежит в основе некоторых современных промышленных способов получения белков, например, антигенов. Недавно промышленным способом были получены некоторые вирусные векторы и лекарственные белки, в настоящее время они проходят клинические и доклинические испытания[270].
В материаловедении и нанотехнологиях[править | править код]
Современные направления в нанотехнологиях обещают принести значительно более разностороннее применение вирусам. С точки зрения материаловедов, вирусы можно рассматривать как органические наночастицы. Их поверхность несёт специальные приспособления для преодоления биологических барьеров клетки-хозяина. Точно определены форма и размер вирусов, а также количество и природа функциональных групп на их поверхности. По существу, вирусы часто используют в материаловедении как «подмости» для ковалентно связанных поверхностных модификаций. Одно из примечательных качеств вирусов — то, что они специально «подогнаны» направленной эволюцией под клетки, выступающие хозяевами. Мощные методы, разработанные биологами, легли в основу инженерных приёмов в наноматериалах, открыв тем самым широкую сферу применения вирусов, выходящую далеко за пределы биологии и медицины[271].
Из-за своих размеров, формы и хорошо изученной химической структуры вирусы использовали как шаблоны для организации материалов на наноуровне. Примером такой недавней работы могут служить исследования, проведённые Исследовательской лабораторией Наваля в Вашингтоне (округ Колумбия) с использованием вируса мозаики коровьего гороха (англ. Cowpea Mosaic Virus (CPMV)) для усиления сигналов в сенсорах с ДНК-микрочипами. В данном случае вирусные частицы разделяли частицы флуоресцентных красителей, которые использовались для передачи сигнала, предотвращая, таким образом, скопление нефлуоресцентных димеров, выступающих как гасители сигнала[272]. Другим примером использования CPMV является применение его как наноразмерного образца для молекулярной электроники[273].
Искусственные вирусы[править | править код]
Многие вирусы могут быть получены de novo, то есть с нуля, а первый искусственный вирус был получен в 2002 году[37]. Несмотря на некоторые неправильные трактовки, при этом синтезируется не сам вирус как таковой, а его геномная ДНК (в случае ДНК-вирусов) или комплементарная копия ДНК его генома (в случае РНК-вирусов). У вирусов многих семейств искусственная ДНК или РНК (последняя получается путём обратной транскрипции синтетической комплементарной ДНК), будучи введённой в клетку, проявляет инфекционные свойства. Иными словами, они содержат всю необходимую информацию для образования новых вирусов. Эту технологию в настоящее время используют для разработки вакцин нового типа[274]. Возможность создавать искусственные вирусы имеет далеко идущие последствия, поскольку вирус не может вымереть, пока известна его геномная последовательность и имеются чувствительные к нему клетки. В наши дни полные геномные последовательности 2408 различных вирусов (в том числе оспы) находятся в публичном доступе в онлайн-базе данных, поддерживаемой Национальными институтами здравоохранения США[275].
Вирусы как оружие[править | править код]
Способность вирусов вызывать опустошительные эпидемии среди людей порождает беспокойство, что вирусы могут использоваться как биологическое оружие. Дополнительные опасения вызвало успешное воссоздание вредоносного вируса испанского гриппа в лаборатории[276]. Другим примером может служить вирус оспы. Он на всём протяжении истории опустошал множество стран вплоть до его окончательного искоренения. Официально образцы вируса оспы хранятся лишь в двух местах в мире — в двух лабораториях в России и США[277]. Опасения, что он может быть использован как оружие, не совсем беспочвенны[277]; вакцина против оспы иногда имеет тяжёлые побочные эффекты — в последние годы до официально объявленного искоренения вируса больше людей серьёзно заболели из-за вакцины, чем от вируса[278], поэтому вакцинация против оспы больше не практикуется повсеместно[279]. По этой причине большая часть современного населения Земли практически не имеет устойчивости к оспе[277].
В массовой культуре[править | править код]
В фильмах и других произведениях мир инфекционных заболеваний, в том числе вирусных, редко представляется достоверно. Исключая фильмы-биографии учёных и фильмы, повествующие о великих эпидемиях прошлого, в большинстве из них центральным событием является вспышка неизвестного болезнетворного агента, появление которого стало результатом акта биотерроризма, инцидента в лаборатории, или же он попал из космоса[280].
В литературе[править | править код]
Вирусная инфекция заложена в основу следующих произведений (список неполный):
- Кодзи Судзуки. «Звонок».
- Кир Булычёв. «Лиловый шар».
- Стивен Кинг. «Противостояние».
- Майкл Крайтон. «Штамм „Андромеда“»[280].
- Джек Лондон. «Алая чума».
- Дэн Браун. «Инферно».
В кинематографе[править | править код]
Вспышка необычной вирусной инфекции лежит в основе сюжета следующих художественных фильмов и сериалов[280]:
- «В город пришла беда» (1966)
- «Часы Пандоры»[en] (1996)
- «28 дней спустя» (англ. 28 Days Later) (2003)
- «28 недель спустя» (англ. 28 Weeks Later) (2007)
- Штамм «Андромеда». Этот фильм, снятый по одноимённой повести Майкла Крайтона, можно назвать наиболее точным в научном плане[280].
- «12 обезьян» (1995)
- «Обитель зла» (2002) и его продолжения.
- «Эпидемия» (1995)
- «Лиловый шар» (1987)
- «Носители» (2009)
- «Я — Легенда» (2007)
- «Заражение» (2011)
- «Карантин» (2008)
- «Карантин 2: Терминал» (2011)
- «Регенезис» (сериал, 2004—2008)
- «Выжить после» (сериал 2013)
- «Спираль» (сериал, 2014—2015)
- «Штамм» (сериал, 2014—2015)
- «Последний корабль» (сериал, 2014—2015)
- «Закрытая школа» (сериал, 2011—2012)
- «Война миров Z» (2013)
- «Инферно» (2016)
- «Эпидемия» (2019)
В мультипликации[править | править код]
В последние годы вирусы нередко становятся «героями» мультфильмов и мультсериалов, среди которых следует назвать, например, «Осмосис Джонс» (США, 2001), «Оззи и Дрикс» (США, 2002—2004 гг.) и «Вирус атакует» (Италия, 2011).
См. также[править | править код]
- Компьютерный вирус
- Медиавирус
- Вироид
- Вирусоподобные частицы
Примечания[править | править код]
- Комментарии
- ↑ На английском языке. В латинском языке вопрос о множественном числе данного слова является спорным. Слово лат. virus принадлежит редкой разновидности II склонения, словам среднего рода на -us: Nom.Acc.Voc. virus, Gen. viri, Dat.Abl. viro. Так же склоняются лат. vulgus и pelagus; в классической латыни множественное число зафиксировано только у последнего: pelage, форма древнегреческого происхождения, где η<εα.
- ↑ 1 2 На данный момент устоявшегося русскоязычного термина, соответствующего англ. realm в таксономии, нет.
- ↑ В различных источниках такое свойство вирусов, как инфекционность, имеет различное значение. Некоторые, например, Большая Советская энциклопедия, определяют вирусы как неклеточные организмы, обладающие свойством вызывать инфекционные болезни у клеточных организмов. Другие, например, Большой Энциклопедический словарь и Биологический энциклопедический словарь, не относят инфекционность к определяющим свойствам вирусов.
- ↑ Как отмечается там, «вид вирусов представляет собой политетический класс вирусов, которые вместе образуют единую линию поколений и занимают особенную экологическую нишу». «Политетический» класс — это таксономическая группа, члены которой имеют несколько общих свойств, хотя и не обязательно имеют все одинаковые свойства. Этим вид вирусов отличается от вышестоящих вирусных таксонов, которые являются «универсальными» классами и имеют набор свойств, обязательных для каждого их члена.
- Использованная литература и источники
- ↑ 1 2 Таксономия вирусов (англ.) на сайте Международного комитета по таксономии вирусов (ICTV). (Дата обращения: 14 мая 2021).
- ↑ 1 2 Koonin E. V., Senkevich T. G., Dolja V. V. The ancient Virus World and evolution of cells // Biol. Direct. — 2006. — Т. 1. — С. 29. — doi:10.1186/1745-6150-1-29. — PMID 16984643. — PMC 1594570.
- ↑ Дмитрий Гапон. «Фильтрующиеся вирусы» открытие в гранях времени // Наука и жизнь. — 2015. — № 6. — С. 38—50. Архивировано 29 июля 2017 года.
- ↑ 1 2 Дмитрий Гапон. «Фильтрующиеся вирусы» открытие в гранях времени // Наука и жизнь. — 2015. — № 7. — С. 31—41. Архивировано 30 июля 2017 года.
- ↑ 1 2 3 4 Dimmock, 2007, p. 4.
- ↑ 1 2 3 Dimmock, 2007, p. 49.
- ↑ 1 2 How many viruses on Earth? Дата обращения: 14 марта 2017. Архивировано 21 марта 2020 года.
- ↑ 1 2 Lawrence C. M., Menon S., Eilers B. J., et al. Structural and functional studies of archaeal viruses (англ.) // J. Biol. Chem. : journal. — 2009. — Vol. 284, no. 19. — P. 12599—12603. — doi:10.1074/jbc.R800078200. — PMID 19158076. — PMC 2675988. Архивировано 24 июля 2018 года.
- ↑ Edwards R. A., Rohwer F. Viral metagenomics // Nat. Rev. Microbiol. — 2005. — Т. 3, № 6. — С. 504—510. — doi:10.1038/nrmicro1163. — PMID 15886693.
- ↑ Casjens S. Desk Encyclopedia of General Virology / Mahy B. W. J. and Van Regenmortel M. H. V.. — Boston: Academic Press, 2010. — С. 167. — ISBN 0-12-375146-2.
- ↑ Русский перевод слова — согласно следующему изданию:
И. Х. Дворецкий. Латинско-русский словарь. Ок. 50000 слов. — Издание второе, переработанное и дополненное. — М.: Русский язык, 1976. — С. [1084] (стб. 1). — 1096 с. — 65 000 экз. Архивная копия от 19 января 2015 на Wayback Machine
virus, i n 1) слизь (cochlearum PM); слизистый сок (pastinanceae PM); семя животных V, PM; 2) ядовитое выделение, яд (serpentus V): ferro v. inest O стрела отравлена; 3) ядовитость, язвительность, жёлочность, едкость (acerbitatis C; linguae, mentis Sil); 4) отвратительный запах, зловоние (paludis Col; animae ursi pestilens v. PM); 5) острый вкус, острота (vini PM); едкость, горечь (sc. maris Lcr; ponti Man).
- ↑ Harper D. virus. The Online Etymology Dictionary (2011). Дата обращения: 23 декабря 2011. Архивировано 19 января 2013 года.
- ↑ Virus // Merriam-Webster.com Dictionary : [англ.] : [арх. 2 ноября 2021]. — Merriam-Webster.
- ↑ William C. Summers. Inventing Viruses (англ.) // Annual Review of Virology. — 2014. — 3 November (vol. 1, iss. 1). — P. 25–35. — ISSN 2327-056X. — doi:10.1146/annurev-virology-031413-085432. Архивировано 7 апреля 2020 года.
- ↑ Harper D. virion. The Online Etymology Dictionary (2011). Дата обращения: 24 декабря 2011. Архивировано 19 января 2013 года.
- ↑ Bordenave G. Louis Pasteur (1822—1895) // Microbes and Infection / Institut Pasteur. — 2003. — Т. 5, № 6. — С. 553—560. — doi:10.1016/S1286-4579(03)00075-3. — PMID 12758285.
- ↑ Shors, 2008, pp. 76—77.
- ↑ 1 2 3 Collier, 1998, p. 3.
- ↑ Гапон Д. «Фильтрующиеся вирусы». Открытие в гранях времени // Наука и жизнь. — 2015. — № 6. — С. 38—50.
- ↑ Dimmock, 2007, p. 4—5.
- ↑ 1 2 Fenner F. Desk Encyclopedia of General Virology / Mahy B. W. J. and Van Regenmortal M. H. V.. — 1. — Oxford, UK: Academic Press, 2009. — С. 15. — ISBN 0-12-375146-2.
- ↑ Shors, 2008, p. 589.
- ↑ D’Herelle F. On an invisible microbe antagonistic toward dysenteric bacilli: brief note by Mr. F. D’Herelle, presented by Mr. Roux (англ.) // Research in Microbiology : journal. — 2007. — Vol. 158, no. 7. — P. 553—554. — doi:10.1016/j.resmic.2007.07.005. — PMID 17855060.
- ↑ Steinhardt E., Israeli C., Lambert R. A. Studies on the cultivation of the virus of vaccinia (англ.) // J. Inf Dis. : journal. — 1913. — Vol. 13, no. 2. — P. 294—300. — doi:10.1093/infdis/13.2.294.
- ↑ Collier, 1998, p. 4.
- ↑ Goodpasture E. W., Woodruff A. M., Buddingh G. J. The cultivation of vaccine and other viruses in the chorioallantoic membrane of chick embryos (англ.) // Science : journal. — 1931. — Vol. 74, no. 1919. — P. 371—372. — doi:10.1126/science.74.1919.371. — PMID 17810781.
- ↑ Rosen, F. S. Isolation of poliovirus—John Enders and the Nobel Prize (англ.) // New England Journal of Medicine : journal. — 2004. — Vol. 351, no. 15. — P. 1481—1483. — doi:10.1056/NEJMp048202. — PMID 15470207.
- ↑ From Nobel Lectures, Physics 1981—1990, (1993) Editor-in-Charge Tore Frängsmyr, Editor Gösta Ekspång, World Scientific Publishing Co., Singapore.
- В 1887 году Бьюст увидел один из крупнейших вирусов, вирус коровьей оспы, в оптический микроскоп, предварительно окрасив его. В то время не было известно, что это вирус. (Buist J. B. Vaccinia and Variola: a study of their life history Churchill, London)
- ↑ Stanley W. M., Loring H. S. The isolation of crystalline tobacco mosaic virus protein from diseased tomato plants (англ.) // Science : journal. — 1936. — Vol. 83, no. 2143. — P. 85. — doi:10.1126/science.83.2143.85. — PMID 17756690.
- ↑ Stanley W. M., Lauffer M. A. Disintegration of tobacco mosaic virus in urea solutions (англ.) // Science : journal. — 1939. — Vol. 89, no. 2311. — P. 345—347. — doi:10.1126/science.89.2311.345. — PMID 17788438.
- ↑ Creager A. N., Morgan G. J. After the double helix: Rosalind Franklin’s research on Tobacco mosaic virus (англ.) // Isis : journal. — 2008. — Vol. 99, no. 2. — P. 239—272. — doi:10.1086/588626. — PMID 18702397. Архивировано 18 августа 2016 года.
- ↑ Dimmock, 2007, p. 12.
- ↑ Norrby E. Nobel Prizes and the emerging virus concept // Arch. Virol.. — 2008. — Т. 153, № 6. — С. 1109—1123. — doi:10.1007/s00705-008-0088-8. — PMID 18446425.
- ↑ Collier, 1998, p. 745.
- ↑ 1 2 Temin H. M., Baltimore D. RNA-directed DNA synthesis and RNA tumor viruses // Adv. Virus Res.. — 1972. — Т. 17. — С. 129—186. — doi:10.1016/S0065-3527(08)60749-6. — PMID 4348509. Архивировано 6 ноября 2015 года.
- ↑ Barré-Sinoussi, F. et al. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS) (англ.) // Science : journal. — 1983. — Vol. 220, no. 4599. — P. 868—871. — doi:10.1126/science.6189183. — PMID 6189183. Архивировано 8 сентября 2015 года.
- ↑ 1 2 Cello J., Paul A. V., Wimmer E. Chemical synthesis of poliovirus cDNA: generation of infectious virus in the absence of natural template (англ.) // Science : journal. — 2002. — Vol. 297, no. 5583. — P. 1016—1018. — doi:10.1126/science.1072266. — PMID 12114528.
- ↑ 1 2 Canchaya C., Fournous G., Chibani-Chennoufi S., Dillmann M. L., Brüssow H. Phage as agents of lateral gene transfer // Curr. Opin. Microbiol.. — 2003. — Т. 6, № 4. — С. 417—424. — doi:10.1016/S1369-5274(03)00086-9. — PMID 12941415.
- ↑ Iyer L. M., Balaji S., Koonin E. V., Aravind L. Evolutionary genomics of nucleo-cytoplasmic large DNA viruses (англ.) // Virus Res. : journal. — 2006. — Vol. 117, no. 1. — P. 156—184. — doi:10.1016/j.virusres.2006.01.009. — PMID 16494962. Архивировано 10 августа 2017 года.
- ↑ 1 2 Sanjuán R., Nebot M. R., Chirico N., Mansky L. M., Belshaw R. Viral mutation rates // Journal of Virology. — 2010. — Октябрь (т. 84, № 19). — С. 9733—9748. — doi:10.1128/JVI.00694-10. — PMID 20660197. — PMC 2937809. Архивировано 8 августа 2014 года.
- ↑ Shors, 2008, pp. 14—16.
- ↑ Collier, 1998, pp. 11—21.
- ↑ Dimmock, 2007, p. 16.
- ↑ Collier, 1998, p. 11.
- ↑ 1 2 3 Mahy W. J. & Van Regenmortel M. H. V. (eds). Desk Encyclopedia of General Virology. — Oxford: Academic Press, 2009. — С. 24. — ISBN 0-12-375146-2.
- ↑ Shors, 2008, p. 574.
- ↑ McClintock, B. The origin and behavior of mutable loci in maize // Proc Natl Acad Sci U S A.. — 1950. — Т. 36, № 6. — С. 344—355. — doi:10.1073/pnas.36.6.344. — PMID 15430309. — PMC 1063197.
- ↑ Collier, 1998, pp. 11—12.
- ↑ Dimmock, 2007, p. 55.
- ↑ Shors, 2008, pp. 551—553.
- ↑ Tsagris E. M., de Alba A. E., Gozmanova M., Kalantidis K. Viroids // Cell. Microbiol.. — 2008. — Т. 10, № 11. — С. 2168. — doi:10.1111/j.1462-5822.2008.01231.x. — PMID 18764915.
- ↑ Shors, 2008, pp. 492—493.
- ↑ 1 2 La Scola B., Desnues C., Pagnier I., Robert C., Barrassi L., Fournous G., Merchat M., Suzan-Monti M., Forterre P., Koonin E., Raoult D. The virophage as a unique parasite of the giant mimivirus (англ.) // Nature : journal. — 2008. — Vol. 455, no. 7209. — P. 100—104. — doi:10.1038/nature07218. — PMID 18690211.
- ↑ Collier, 1998, p. 777.
- ↑ Dimmock, 2007, pp. 55—57.
- ↑ 1 2 Mahy W. J. & Van Regenmortel M. H. V. (eds). Desk Encyclopedia of General Virology. — Oxford: Academic Press, 2009. — С. 28. — ISBN 0-12-375146-2.
- ↑ 1 2 Mahy W. J. & Van Regenmortel M. H. V. (eds). Desk Encyclopedia of General Virology. — Oxford: Academic Press, 2009. — С. 26. — ISBN 0-12-375146-2.
- ↑ Dimmock, 2007, pp. 15—16.
- ↑ Liberski P. P. Prion diseases: a riddle wrapped in a mystery inside an enigma (англ.) // Folia Neuropathol : journal. — 2008. — Vol. 46, no. 2. — P. 93—116. — PMID 18587704.
- ↑ Инге-Вечтомов С. Г. Генетика с основами селекции. — СПб.: Издательство Н-Л, 2010. — С. 298. — 718 с. — ISBN 978-5-94869-105-3.
- ↑ Belay E. D. and Schonberger L. B. Desk Encyclopedia of Human and Medical Virology (англ.). — Boston: Academic Press, 2009. — P. 497—504. — ISBN 0-12-375147-0.
- ↑ Lupi O., Dadalti P., Cruz E., Goodheart C. Did the first virus self-assemble from self-replicating prion proteins and RNA? (англ.) // Med. Hypotheses : journal. — 2007. — Vol. 69, no. 4. — P. 724—730. — doi:10.1016/j.mehy.2007.03.031. — PMID 17512677.
- ↑ Holmes E. C. Viral evolution in the genomic age // PLoS Biol.. — 2007. — Т. 5, № 10. — С. e278. — doi:10.1371/journal.pbio.0050278. — PMID 17914905. — PMC 1994994. Архивировано 5 ноября 2013 года.
- ↑ Wimmer E., Mueller S., Tumpey T. M., Taubenberger J. K. Synthetic viruses: a new opportunity to understand and prevent viral disease (англ.) // Nature Biotechnology : journal. — Nature Publishing Group, 2009. — Vol. 27, no. 12. — P. 1163—1172. — doi:10.1038/nbt.1593. — PMID 20010599. — PMC 2819212.
- ↑ Horn M. Chlamydiae as symbionts in eukaryotes // Annual Review of Microbiology. — 2008. — Т. 62. — С. 113—131. — doi:10.1146/annurev.micro.62.081307.162818. — PMID 18473699. Архивировано 11 ноября 2013 года.
- ↑ Ammerman N. C., Beier-Sexton M., Azad A. F. Laboratory maintenance of Rickettsia rickettsii // Current Protocols (англ.) (рус. in Microbiology. — 2008. — Т. Chapter 3. — С. Unit 3A.5. — doi:10.1002/9780471729259.mc03a05s11. — PMID 19016440. — PMC 2725428.
- ↑ Seed K. D., Lazinski D. W., Calderwood S. B., Camilli A. A bacteriophage encodes its own CRISPR/Cas adaptive response to evade host innate immunity (англ.) // Nature : journal. — 2013. — Vol. 494, no. 7438. — P. 489—491. — doi:10.1038/nature11927. — PMID 23446421. — PMC 3587790. Архивировано 5 марта 2016 года.
- ↑ 1 2 Collier, 1998, pp. 33—55.
- ↑ Stefan Sirucek. Ancient “Giant Virus” Revived From Siberian Permafrost, National Geographic (3 марта 2014). Архивировано 25 июня 2018 года. Дата обращения: 3 марта 2014.
- ↑ Collier, 1998, pp. 33—37.
- ↑ Kiselev N. A., Sherman M. B., Tsuprun V. L. Negative staining of proteins // Electron Microsc. Rev.. — 1990. — Т. 3, № 1. — С. 43—72. — doi:10.1016/0892-0354(90)90013-I. — PMID 1715774.
- ↑ Collier, 1998, p. 40.
- ↑ Caspar D. L., Klug A. Physical principles in the construction of regular viruses (англ.) // Cold Spring Harb. Symp. Quant. Biol. : journal. — 1962. — Vol. 27. — P. 1—24. — PMID 14019094. Архивировано 5 марта 2016 года.
- ↑ Crick F. H., Watson J. D. Structure of small viruses (англ.) // Nature. — 1956. — Vol. 177, no. 4506. — P. 473—475. — doi:10.1038/177473a0. — PMID 13309339. Архивировано 5 июля 2013 года.
- ↑ Falvo, M.R.; S. Washburn, R. Superfine, M. Finch, F. P. Brooks Jr, V. Chi, R. M. Taylor 2nd. Manipulation of individual viruses: friction and mechanical properties (англ.) // Biophysical Journal : journal. — 1997. — Vol. 72, no. 3. — P. 1396—1403. — doi:10.1016/S0006-3495(97)78786-1. — PMID 9138585. — PMC 1184522.
- ↑ Kuznetsov, Yu. G.; A. J. Malkin, R. W. Lucas, M. Plomp, A. McPherson. Imaging of viruses by atomic force microscopy // J Gen Virol. — 2001. — Т. 82, № 9. — С. 2025—2034. — PMID 11514711. (недоступная ссылка)
- ↑ Collier, 1998, p. 37.
- ↑ Collier, 1998, pp. 40, 42.
- ↑ Casens, S. Desk Encyclopedia of General Virology. — Boston: Academic Press, 2009. — С. 167—174. — ISBN 0-12-375146-2.
- ↑ Rossmann M. G., Mesyanzhinov V. V., Arisaka F., Leiman P. G. The bacteriophage T4 DNA injection machine (англ.) // Curr. Opin. Struct. Biol.. — 2004. — Vol. 14, no. 2. — P. 171—180. — doi:10.1016/j.sbi.2004.02.001. — PMID 15093831.
- ↑ Collier, 1998, pp. 42—43.
- ↑ Long G. W., Nobel J., Murphy F. A., Herrmann K. L., Lourie B. Experience with electron microscopy in the differential diagnosis of smallpox (англ.) // Appl Microbiol : journal. — 1970. — Vol. 20, no. 3. — P. 497—504. — PMID 4322005. — PMC 376966.
- ↑ Xiao C., Kuznetsov Y. G., Sun S., Hafenstein S. L., Kostyuchenko V. A., Chipman P. R., Suzan-Monti M., Raoult D., McPherson A., Rossmann M. G. Structural studies of the giant mimivirus // PLoS Biol. — 2009. — Т. 7, вып. 4. — С. e92. — doi:10.1371/journal.pbio.1000092. — PMID 19402750. Архивировано 5 июля 2020 года.
- ↑ Klose T., Kuznetsov Y. G., Xiao C., Sun S., McPherson A., Rossmann M. G. The three-dimensional structure of Mimivirus // Intervirology. — 2010. — Т. 53, вып. 5. — С. 268—273. — doi:10.1159/000312911. — PMID 20551678. Архивировано 5 июня 2022 года.
- ↑ World’s biggest virus discovered in ocean depths near Chile. Дата обращения: 12 октября 2011. Архивировано 1 февраля 2013 года.
- ↑ 1 2 3 Collier, 1998, pp. 96—99.
- ↑ Saunders, Venetia A.; Carter, John. Virology: principles and applications. — Chichester: John Wiley & Sons, 2007. — С. 72. — ISBN 0-470-02387-2.
- ↑ Van Etten J. L., Lane L. C., Dunigan D. D. DNA viruses: the really big ones (giruses) // Annual Review of Microbiology. — 2010. — Т. 64. — С. 83—99. — doi:10.1146/annurev.micro.112408.134338. — PMID 20690825. — PMC 2936810. Архивировано 30 сентября 2021 года.
- ↑ Pressing J., Reanney D. C. Divided genomes and intrinsic noise // J Mol Evol. — 1984. — Т. 20, № 2. — С. 135—146. — doi:10.1007/BF02257374. — PMID 6433032.
- ↑ Duffy S., Holmes E. C. Validation of high rates of nucleotide substitution in geminiviruses: phylogenetic evidence from East African cassava mosaic viruses (англ.) // Journal of General Virology (англ.) (рус. : journal. — Microbiology Society (англ.) (рус., 2009. — Vol. 90, no. Pt 6. — P. 1539—1547. — doi:10.1099/vir.0.009266-0. — PMID 19264617. (недоступная ссылка)
- ↑ Pan X. P., Li L. J., Du W. B., Li M. W., Cao H. C., Sheng J. F. Differences of YMDD mutational patterns, precore/core promoter mutations, serum HBV DNA levels in lamivudine-resistant hepatitis B genotypes B and C (англ.) // J. Viral Hepat. : journal. — 2007. — Vol. 14, no. 11. — P. 767—774. — doi:10.1111/j.1365-2893.2007.00869.x. — PMID 17927612. Архивировано 18 марта 2021 года.
- ↑ Hampson A. W., Mackenzie J. S. The influenza viruses // Med. J. Aust.. — 2006. — Т. 185, № 10 Suppl. — С. S39—43. — PMID 17115950. Архивировано 16 мая 2013 года.
- ↑ Metzner K. J. Detection and significance of minority quasispecies of drug-resistant HIV-1 (англ.) // J HIV Ther : journal. — 2006. — Vol. 11, no. 4. — P. 74—81. — PMID 17578210.
- ↑ Goudsmit, Jaap. Viral Sex. Oxford Univ Press, 1998. ISBN 978-0-19-512496-5, ISBN 0-19-512496-0.
- ↑ Worobey M., Holmes E. C. Evolutionary aspects of recombination in RNA viruses (англ.) // Journal of General Virology (англ.) (рус. : journal. — Microbiology Society (англ.) (рус., 1999. — Vol. 80 (Pt 10). — P. 2535—2543. — PMID 10573145. (недоступная ссылка)
- ↑ Lukashev A. N. Role of recombination in evolution of enteroviruses (англ.) // Rev. Med. Virol. : journal. — 2005. — Vol. 15, no. 3. — P. 157—167. — doi:10.1002/rmv.457. — PMID 15578739.
- ↑ Umene K. Mechanism and application of genetic recombination in herpesviruses (англ.) // Rev. Med. Virol. : journal. — 1999. — Vol. 9, no. 3. — P. 171—182. — doi:10.1002/(SICI)1099-1654(199907/09)9:3<171::AID-RMV243>3.0.CO;2-A. — PMID 10479778.
- ↑ Collier, 1998, pp. 75—91.
- ↑ Dimmock, 2007, p. 70.
- ↑ Boevink P., Oparka K. J. Virus-host interactions during movement processes // Plant Physiol.. — 2005. — Т. 138, № 4. — С. 1815—1821. — doi:10.1104/pp.105.066761. — PMID 16172094. — PMC 1183373.
- ↑ Dimmock, 2007, p. 71.
- ↑ Barman S., Ali A., Hui E. K., Adhikary L., Nayak D. P. Transport of viral proteins to the apical membranes and interaction of matrix protein with glycoproteins in the assembly of influenza viruses (англ.) // Virus Res. : journal. — 2001. — Vol. 77, no. 1. — P. 61—69. — doi:10.1016/S0168-1702(01)00266-0. — PMID 11451488.
- ↑ Shors, 2008, pp. 60, 597.
- ↑ Dimmock, 2007, Chapter 15, Mechanisms in virus latentcy, pp. 243—259.
- ↑ Dimmock, 2007, pp. 185—187.
- ↑ Collier, 1998, p. 78.
- ↑ Shors, 2008, p. 54.
- ↑ Collier, 1998, p. 79.
- ↑ Collier, 1998, pp. 88—89.
- ↑ Staginnus C., Richert-Pöggeler K. R. Endogenous pararetroviruses: two-faced travelers in the plant genome (англ.) // Trends in Plant Science : journal. — Cell Press, 2006. — Vol. 11, no. 10. — P. 485—491. — doi:10.1016/j.tplants.2006.08.008. — PMID 16949329.
- ↑ Collier, 1998, pp. 115—146.
- ↑ Collier, 1998, p. 115.
- ↑ Roulston A., Marcellus R. C., Branton P. E. Viruses and apoptosis // Annu. Rev. Microbiol.. — 1999. — Т. 53. — С. 577—628. — doi:10.1146/annurev.micro.53.1.577. — PMID 10547702.
- ↑ Alwine J. C. Modulation of host cell stress responses by human cytomegalovirus (англ.) // Curr. Top. Microbiol. Immunol. : journal. — 2008. — Vol. 325. — P. 263—279. — doi:10.1007/978-3-540-77349-8_15. — PMID 18637511.
- ↑ Sinclair J. Human cytomegalovirus: Latency and reactivation in the myeloid lineage (англ.) // J. Clin. Virol. : journal. — 2008. — Vol. 41, no. 3. — P. 180—185. — doi:10.1016/j.jcv.2007.11.014. — PMID 18164651.
- ↑ Jordan M. C., Jordan G. W., Stevens J. G., Miller G. Latent herpesviruses of humans // Ann. Intern. Med.. — 1984. — Т. 100, № 6. — С. 866—880. — PMID 6326635.
- ↑ Sissons J. G., Bain M., Wills M. R. Latency and reactivation of human cytomegalovirus // J. Infect.. — 2002. — Т. 44, № 2. — С. 73—77. — doi:10.1053/jinf.2001.0948. — PMID 12076064.
- ↑ Barozzi P., Potenza L., Riva G., Vallerini D., Quadrelli C., Bosco R., Forghieri F., Torelli G., Luppi M. B cells and herpesviruses: a model of lymphoproliferation (англ.) // Autoimmun Rev : journal. — 2007. — Vol. 7, no. 2. — P. 132—136. — doi:10.1016/j.autrev.2007.02.018. — PMID 18035323.
- ↑ Subramanya D., Grivas P. D. HPV and cervical cancer: updates on an established relationship (англ.) // Postgrad Med : journal. — 2008. — Vol. 120, no. 4. — P. 7—13. — doi:10.3810/pgm.2008.11.1928. — PMID 19020360.
- ↑ Crawford, Dorothy H. Viruses: A Very Short Introduction (англ.). — Oxford University Press, 2011. — P. 16. — ISBN 0-19-957485-5.
- ↑ Shors, 2008, p. 388.
- ↑ Shors, 2008, p. 353.
- ↑ Dimmock, 2007, p. 272.
- ↑ Baggesen D. L., Sørensen G., Nielsen E. M., Wegener H. C. Phage typing of Salmonella Typhimurium – is it still a useful tool for surveillance and outbreak investigation? (англ.) // Eurosurveillance (англ.) (рус. : journal. — 2010. — Vol. 15, no. 4. — P. 19471. — PMID 20122382. Архивировано 17 февраля 2013 года.
- ↑ Shors, 2008, pp. 49—50.
- ↑ Systema Naturae 2000: Overview. Дата обращения: 17 марта 2013. Архивировано 26 августа 2005 года.
- ↑ Taxonomicon: Aphanobionta. Дата обращения: 17 марта 2013. Архивировано 14 марта 2022 года.
- ↑ Taxonomicon: Acytota. Дата обращения: 17 марта 2013. Архивировано 7 марта 2016 года.
- ↑ Систематика органического мира: Бесклеточные (Acellularia). Дата обращения: 17 марта 2013. Архивировано из оригинала 23 февраля 2012 года.
- ↑ NCBI taxonomy. Дата обращения: 29 сентября 2017. Архивировано 6 апреля 2018 года.
- ↑ Lwoff A., Horne R. W., Tournier P. A virus system (фр.) // C. R. Hebd. Seances Acad. Sci.. — 1962. — Vol. 254. — P. 4225—4227. — PMID 14467544.
- ↑ Lwoff A., Horne R., Tournier P. A system of viruses // Cold Spring Harb. Symp. Quant. Biol.. — 1962. — Т. 27. — С. 51—55. — PMID 13931895.
- ↑ 1 2 3 Wolf Y. I., Kazlauskas D., Iranzo J., Lucía-Sanz A., Kuhn J. H., Krupovic M., Dolja V. V., Koonin E. V. Origins and Evolution of the Global RNA Virome. (англ.) // MBio. — 2018. — 27 November (vol. 9, no. 6). — doi:10.1128/mBio.02329-18. — PMID 30482837. [исправить]
- ↑ 1 2 3 4 5 Koonin E. V., Krupovic M., Agol V. I. The Baltimore Classification of Viruses 50 Years Later: How Does It Stand in the Light of Virus Evolution? (англ.) // Microbiology And Molecular Biology Reviews : MMBR. — 2021. — 18 August (vol. 85, no. 3). — P. e0005321—0005321. — doi:10.1128/MMBR.00053-21. — PMID 34259570. [исправить]
- ↑ Knipe, 2007, p. 27.
- ↑ Delwart E. L. Viral metagenomics (англ.) // Rev. Med. Virol.. — 2007. — Vol. 17, no. 2. — P. 115—131. — doi:10.1002/rmv.532. — PMID 17295196.
- ↑ ICVC&N, 2018, 3.2.
- ↑ ICVC&N, 2018, 3.23.
- ↑ ICVC&N, 2018, 3.3.
- ↑ King A. M. Q., Lefkowitz E., Adams M. J., Carstens E. B. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses (англ.). — Elsevier, 2011. — P. 6. — ISBN 0-12-384684-6.
- ↑ Koonin EV, Dolja VV, Krupovic M, Varsani A, Wolf YI, Yutin N, Zerbini M, Kuhn JH. Create a megataxonomic framework, filling all principal taxonomic ranks, for DNA viruses encoding vertical jelly roll-type major capsid proteins (англ.) (docx). International Committee on Taxonomy of Viruses (18 октября 2019). Дата обращения: 10 июня 2020. Архивировано 4 января 2022 года.
- ↑ Koonin, Eugene & Dolja, Valerian & Krupovic, Mart & Varsani, Arvind & Wolf, Yuri & Yutin, Natalya & Zerbini, Francisco & Kuhn, Jens. Create a megataxonomic framework, filling all principal/primary taxonomic ranks, for dsDNA viruses encoding HK97-type major capsid proteins. — 2019. — doi:10.13140/RG.2.2.16564.19842.
- ↑ 1 2 Baltimore D. The strategy of RNA viruses // Harvey Lect.. — 1974. — Т. 70 Series. — С. 57—74. — PMID 4377923.
- ↑ Mayo M. A. Developments in plant virus taxonomy since the publication of the 6th ICTV Report. International Committee on Taxonomy of Viruses (англ.) // Arch. Virol. : journal. — 1999. — Vol. 144, no. 8. — P. 1659—1666. — doi:10.1007/s007050050620. — PMID 10486120.
- ↑ de Villiers E. M., Fauquet C., Broker T. R., Bernard H. U., zur Hausen H. Classification of papillomaviruses // Virology. — 2004. — Т. 324, № 1. — С. 17—27. — doi:10.1016/j.virol.2004.03.033. — PMID 15183049.
- ↑ Temin H. M., Baltimore D. RNA-directed DNA synthesis and RNA tumor viruses // Adv. Virus Res.. — 1972. — Т. 17. — С. 129—186. — PMID 4348509.
- ↑ Lazarowitz SD (2007) «Plant viruses», in «Fields’ Virology», 5th edition, volume 1, pp. 679—683, Wolters Kluwer / Lippincott Williams & Wilkins, ISBN 0-7817-6060-7
- ↑ Seeger C, Zoulin F, Mason WS (2007) «Hepadnaviruses», in «Fields’ Virology», 5th edition, volume 2, pp. 2977-3029, Wolters Kluwer / Lippincott Williams & Wilkins, ISBN 0-7817-6060-7
- ↑ The Big Picture Book of Viruses:Family Groups – The Baltimore Method. Дата обращения: 21 мая 2015. Архивировано 28 апреля 2013 года.
- ↑ Komaroff A. L. Is human herpesvirus-6 a trigger for chronic fatigue syndrome? (англ.) // J. Clin. Virol. : journal. — 2006. — Vol. 37 Suppl 1. — P. S39—46. — doi:10.1016/S1386-6532(06)70010-5. — PMID 17276367.
- ↑ Chen C., Chiu Y., Wei F., Koong F., Liu H., Shaw C., Hwu H., Hsiao K. High seroprevalence of Borna virus infection in schizophrenic patients, family members and mental health workers in Taiwan (англ.) // Mol Psychiatry : journal. — 1999. — Vol. 4, no. 1. — P. 33—38. — doi:10.1038/sj.mp.4000484. — PMID 10089006.
- ↑ Margolis T. P., Elfman F. L., Leib D., et al. Spontaneous reactivation of herpes simplex virus type 1 in latently infected murine sensory ganglia (англ.) // J. Virol. : journal. — 2007. — Vol. 81, no. 20. — P. 11069—11074. — doi:10.1128/JVI.00243-07. — PMID 17686862. — PMC 2045564.
- ↑ Whitley R. J., Roizman B. Herpes simplex virus infections (англ.) // The Lancet. — Elsevier, 2001. — Vol. 357, no. 9267. — P. 1513—1518. — doi:10.1016/S0140-6736(00)04638-9. — PMID 11377626. Архивировано 30 марта 2018 года.
- ↑ Barton E. S., White D. W., Cathelyn J. S., et al. Herpesvirus latency confers symbiotic protection from bacterial infection (англ.) // Nature : journal. — 2007. — Vol. 447, no. 7142. — P. 326—329. — doi:10.1038/nature05762. — PMID 17507983.
- ↑ Bertoletti A., Gehring A. Immune response and tolerance during chronic hepatitis B virus infection (англ.) // Hepatol. Res. : journal. — 2007. — Vol. 37 Suppl 3. — P. S331—S338. — doi:10.1111/j.1872-034X.2007.00221.x. — PMID 17931183.
- ↑ Rodrigues C., Deshmukh M., Jacob T., Nukala R., Menon S., Mehta A. Significance of HBV DNA by PCR over serological markers of HBV in acute and chronic patients (англ.) // Indian journal of medical microbiology : journal. — 2001. — Vol. 19, no. 3. — P. 141—144. — PMID 17664817. Архивировано 8 января 2007 года.
- ↑ Nguyen V. T., McLaws M. L., Dore G. J. Highly endemic hepatitis B infection in rural Vietnam (англ.) // Journal of Gastroenterology and Hepatology : journal. — 2007. — Vol. 22, no. 12. — P. 2093—2100. — doi:10.1111/j.1440-1746.2007.05010.x. — PMID 17645465.
- ↑ Fowler M. G., Lampe M. A., Jamieson D. J., Kourtis A. P., Rogers M. F. Reducing the risk of mother-to-child human immunodeficiency virus transmission: past successes, current progress and challenges, and future directions (англ.) // American Journal of Obstetrics and Gynecology (англ.) (рус. : journal. — Elsevier, 2007. — Vol. 197, no. 3 Suppl. — P. S3—9. — doi:10.1016/j.ajog.2007.06.048. — PMID 17825648.
- ↑ Sauerbrei A., Wutzler P. The congenital varicella syndrome // Journal of perinatology : official journal of the California Perinatal Association. — 2000. — Т. 20, № 8 Pt 1. — С. 548—554. — PMID 11190597.
- ↑ Garnett G. P. Role of herd immunity in determining the effect of vaccines against sexually transmitted disease (англ.) // J. Infect. Dis. : journal. — 2005. — Vol. 191 Suppl 1. — P. S97—106. — doi:10.1086/425271. — PMID 15627236. (недоступная ссылка)
- ↑ Platonov A. E. Влияние погодных условий на эпидемиологию трансмиссивных инфекций (на примере лихорадки Западного Нила в России) // Vestn. Akad. Med. Nauk SSSR. — 2006. — № 2. — С. 25—29. — PMID 16544901.
- ↑ Shors, 2008, p. 198.
- ↑ Shors, 2008, pp. 199, 209.
- ↑ Shors, 2008, p. 19.
- ↑ Shors, 2008, p. 126.
- ↑ 1 2 Shors, 2008, pp. 193—194.
- ↑ Shors, 2008, p. 194.
- ↑ Shors, 2008, pp. 192—193.
- ↑ * Ranlet P. The British, the Indians, and smallpox: what actually happened at Fort Pitt in 1763? (англ.) // Pa Hist : journal. — 2000. — Vol. 67, no. 3. — P. 427—441. — PMID 17216901.
- Van Rijn K. “Lo! The poor Indian!” colonial responses to the 1862–63 smallpox epidemic in British Columbia and Vancouver Island (англ.) // Can Bull Med Hist : journal. — 2006. — Vol. 23, no. 2. — P. 541—560. — PMID 17214129.
- Patterson K. B., Runge T. Smallpox and the Native American // Am. J. Med. Sci.. — 2002. — Т. 323, № 4. — С. 216—222. — doi:10.1097/00000441-200204000-00009. — PMID 12003378.
- Sessa R., Palagiano C., Scifoni M. G., di Pietro M., Del Piano M. The major epidemic infections: a gift from the Old World to the New? (англ.) // Panminerva Med : journal. — 1999. — Vol. 41, no. 1. — P. 78—84. — PMID 10230264.
- Bianchine P. J., Russo T. A. The role of epidemic infectious diseases in the discovery of America (англ.) // Allergy Proc : journal. — 1992. — Vol. 13, no. 5. — P. 225—232. — doi:10.2500/108854192778817040. — PMID 1483570. Архивировано 6 декабря 2012 года.
- Hauptman L. M. Smallpox and American Indian; Depopulation in Colonial New York (англ.) // N Y State J Med : journal. — 1979. — Vol. 79, no. 12. — P. 1945—1949. — PMID 390434.
- Fortuine R. Smallpox decimates the Tlingit (1787) (англ.) // Alaska Med. — 1988. — Vol. 30, no. 3. — P. 109. — PMID 3041871.
- ↑ Collier, 1998, pp. 409—415.
- ↑ Patterson K. D., Pyle G. F. The geography and mortality of the 1918 influenza pandemic (англ.) // Bull Hist Med. : journal. — 1991. — Vol. 65, no. 1. — P. 4—21. — PMID 2021692.
- ↑ Johnson N. P., Mueller J. Updating the accounts: global mortality of the 1918–1920 “Spanish” influenza pandemic (англ.) // Bull Hist Med : journal. — 2002. — Vol. 76, no. 1. — P. 105—115. — doi:10.1353/bhm.2002.0022. — PMID 11875246.
- ↑ Gao F., Bailes E., Robertson D. L., et al. Origin of HIV-1 in the Chimpanzee Pan troglodytes troglodytes (англ.) // Nature : journal. — 1999. — Vol. 397, no. 6718. — P. 436—441. — doi:10.1038/17130. — PMID 9989410. Архивировано 23 февраля 2005 года.
- ↑ Shors, 2008, p. 447.
- ↑ Mawar N., Saha S., Pandit A., Mahajan U. The third phase of HIV pandemic: social consequences of HIV/AIDS stigma & discrimination & future needs (англ.) // Indian J. Med. Res. : journal. — 2005. — Vol. 122, no. 6. — P. 471—484. — PMID 16517997. Архивировано 4 марта 2016 года. Архивированная копия. Дата обращения: 27 февраля 2013. Архивировано из оригинала 4 марта 2016 года.
- ↑ Status of the global HIV epidemic (PDF) (недоступная ссылка — история). UNAIDS (2008). Дата обращения: 15 сентября 2008. Архивировано 9 марта 2013 года.
- ↑ Towner J. S., Khristova M. L., Sealy T. K., et al. Marburgvirus genomics and association with a large hemorrhagic fever outbreak in Angola (англ.) // J. Virol. : journal. — 2006. — Vol. 80, no. 13. — P. 6497—6516. — doi:10.1128/JVI.00069-06. — PMID 16775337. — PMC 1488971. Архивировано 25 июля 2018 года.
- ↑ Einstein M. H., Schiller J. T., Viscidi R. P., Strickler H. D., Coursaget P., Tan T., Halsey N., Jenkins D. Clinician’s guide to human papillomavirus immunology: knowns and unknowns (англ.) // The Lancet : journal. — Elsevier, 2009. — Vol. 9, no. 6. — P. 347—356. — doi:10.1016/S1473-3099(09)70108-2. — PMID 19467474.
- ↑ Shuda M., Feng H., Kwun H. J., Rosen S. T., Gjoerup O., Moore P. S., Chang Y. T antigen mutations are a human tumor-specific signature for Merkel cell polyomavirus (англ.) // Proceedings of the National Academy of Sciences : journal. — United States National Academy of Sciences, 2008. — Vol. 105, no. 42. — P. 16272—16277. — doi:10.1073/pnas.0806526105. — PMID 18812503. — PMC 2551627.
- ↑ Pulitzer M. P., Amin B. D., Busam K. J. Merkel cell carcinoma: review // Advances in Anatomic Pathology. — 2009. — Т. 16, № 3. — С. 135—144. — doi:10.1097/PAP.0b013e3181a12f5a. — PMID 19395876.
- ↑ Koike K. Hepatitis C virus contributes to hepatocarcinogenesis by modulating metabolic and intracellular signalling pathways (англ.) // J. Gastroenterol. Hepatol. : journal. — 2007. — Vol. 22 Suppl 1. — P. S108—S111. — doi:10.1111/j.1440-1746.2006.04669.x. — PMID 17567457.
- ↑ Hu J., Ludgate L. HIV-HBV and HIV-HCV coinfection and liver cancer development (англ.) // Cancer Treat. Res. : journal. — 2007. — Vol. 133. — P. 241—252. — doi:10.1007/978-0-387-46816-7_9. — PMID 17672044.
- ↑ Bellon M., Nicot C. Telomerase: a crucial player in HTLV-I-induced human T-cell leukemia (англ.) // Cancer genomics & proteomics : journal. — 2007. — Vol. 4, no. 1. — P. 21—25. — PMID 17726237.
- ↑ Schiffman M., Castle P. E., Jeronimo J., Rodriguez A. C., Wacholder S. Human papillomavirus and cervical cancer (англ.) // The Lancet. — Elsevier, 2007. — Vol. 370, no. 9590. — P. 890—907. — doi:10.1016/S0140-6736(07)61416-0. — PMID 17826171.
- ↑ Klein E., Kis L. L., Klein G. Epstein-Barr virus infection in humans: from harmless to life endangering virus-lymphocyte interactions (англ.) // Oncogene (англ.) (рус. : journal. — 2007. — Vol. 26, no. 9. — P. 1297—1305. — doi:10.1038/sj.onc.1210240. — PMID 17322915.
- ↑ zur Hausen H. Novel human polyomaviruses—re-emergence of a well known virus family as possible human carcinogens (англ.) // International Journal of Cancer. Journal International Du Cancer : journal. — 2008. — Vol. 123, no. 2. — P. 247—250. — doi:10.1002/ijc.23620. — PMID 18449881.
- ↑ Alberts, Bruce; Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walters. Molecular Biology of the Cell; Fourth Edition (англ.). — New York and London: Garland Science (англ.) (рус., 2002. — ISBN 0-8153-3218-1. Архивная копия от 18 сентября 2009 на Wayback Machine
- ↑ Ding S. W., Voinnet O. Antiviral immunity directed by small RNAs (англ.) // Cell. — Cell Press, 2007. — Vol. 130, no. 3. — P. 413—426. — doi:10.1016/j.cell.2007.07.039. — PMID 17693253. — PMC 2703654.
- ↑ Patton J. T., Vasquez-Del Carpio R., Spencer E. Replication and transcription of the rotavirus genome (англ.) // Curr. Pharm. Des. : journal. — 2004. — Vol. 10, no. 30. — P. 3769—3777. — doi:10.2174/1381612043382620. — PMID 15579070.
- ↑ Jayaram H., Estes M. K., Prasad B. V. Emerging themes in rotavirus cell entry, genome organization, transcription and replication (англ.) // Virus Res. : journal. — 2004. — Vol. 101, no. 1. — P. 67—81. — doi:10.1016/j.virusres.2003.12.007. — PMID 15010218.
- ↑ Greer S., Alexander G. J. Viral serology and detection // Baillieres Clin. Gastroenterol.. — 1995. — Т. 9, № 4. — С. 689—721. — doi:10.1016/0950-3528(95)90057-8. — PMID 8903801.
- ↑ Matter L., Kogelschatz K., Germann D. Serum levels of rubella virus antibodies indicating immunity: response to vaccination of subjects with low or undetectable antibody concentrations (англ.) // J. Infect. Dis. : journal. — 1997. — Vol. 175, no. 4. — P. 749—755. — doi:10.1086/513967. — PMID 9086126.
- ↑ Mallery D. L., McEwan W. A., Bidgood S. R., Towers G. J., Johnson C. M., James L. C. Antibodies mediate intracellular immunity through tripartite motif-containing 21 (TRIM21) (англ.) // Proceedings of the National Academy of Sciences : journal. — United States National Academy of Sciences, 2010. — November (vol. 107, no. 46). — P. 19985—19990. — doi:10.1073/pnas.1014074107. — PMID 21045130. — PMC 2993423.
- ↑ Cascalho M., Platt J. L. Novel functions of B cells // Crit. Rev. Immunol.. — 2007. — Т. 27, № 2. — С. 141—151. — PMID 17725500.
- ↑ Le Page C., Génin P., Baines M. G., Hiscott J. Interferon activation and innate immunity // Rev Immunogenet. — 2000. — Т. 2, № 3. — С. 374—386. — PMID 11256746.
- ↑ Maurice R. Hilleman. Strategies and mechanisms for host and pathogen survival in acute and persistent viral infections (англ.) // Proceedings of the National Academy of Sciences. — National Academy of Sciences, 2004-10-05. — Vol. 101, iss. suppl 2. — P. 14560—14566. — doi:10.1073/pnas.0404758101.
- ↑ Asaria P., MacMahon E. Measles in the United Kingdom: can we eradicate it by 2010? (англ.) // BMJ : journal. — 2006. — Vol. 333, no. 7574. — P. 890—895. — doi:10.1136/bmj.38989.445845.7C. — PMID 17068034. — PMC 1626346. Архивировано 7 марта 2016 года.
- ↑ Lane J. M. Mass vaccination and surveillance/containment in the eradication of smallpox (англ.) // Curr. Top. Microbiol. Immunol. : journal. — 2006. — Vol. 304. — P. 17—29. — doi:10.1007/3-540-36583-4_2. — PMID 16989262.
- ↑ Arvin A. M., Greenberg H. B. New viral vaccines // Virology. — 2006. — Т. 344, № 1. — С. 240—249. — doi:10.1016/j.virol.2005.09.057. — PMID 16364754.
- ↑ Pastoret P. P., Schudel A. A., Lombard M. Conclusions—future trends in veterinary vaccinology (англ.) // Rev. – Off. Int. Epizoot. : journal. — 2007. — Vol. 26, no. 2. — P. 489—494. — PMID 17892169.
- ↑ Palese P. Making better influenza virus vaccines? // Emerging Infect. Dis.. — 2006. — Т. 12, № 1. — С. 61—65. — doi:10.3201/eid1201.051043. — PMID 16494719. Архивировано 2 октября 2012 года.
- ↑ Thomssen R. Live attenuated versus killed virus vaccines // Monographs in allergy. — 1975. — Т. 9. — С. 155—176. — PMID 1090805.
- ↑ McLean A. A. Development of vaccines against hepatitis A and hepatitis B (англ.) // Rev. Infect. Dis. : journal. — 1986. — Vol. 8, no. 4. — P. 591—598. — PMID 3018891.
- ↑ Casswall T. H., Fischler B. Vaccination of the immunocompromised child // Expert review of vaccines. — 2005. — Т. 4, № 5. — С. 725—738. — doi:10.1586/14760584.4.5.725. — PMID 16221073.
- ↑ Barnett E. D., Wilder-Smith A., Wilson M. E. Yellow fever vaccines and international travelers // Expert Rev Vaccines. — 2008. — Т. 7, № 5. — С. 579—587. — doi:10.1586/14760584.7.5.579. — PMID 18564013.
- ↑ Magden J., Kääriäinen L., Ahola T. Inhibitors of virus replication: recent developments and prospects (англ.) // Appl. Microbiol. Biotechnol. : journal. — 2005. — Vol. 66, no. 6. — P. 612—621. — doi:10.1007/s00253-004-1783-3. — PMID 15592828.
- ↑ Mindel A., Sutherland S. Genital herpes — the disease and its treatment including intravenous acyclovir (англ.) // J. Antimicrob. Chemother. : journal. — 1983. — Vol. 12 Suppl B. — P. 51—59. — PMID 6355051.
- ↑ Witthöft T., Möller B., Wiedmann K. H., et al. Safety, tolerability and efficacy of peginterferon alpha-2a and ribavirin in chronic hepatitis C in clinical practice: The German Open Safety Trial (англ.) // J. Viral Hepat. : journal. — 2007. — Vol. 14, no. 11. — P. 788—796. — doi:10.1111/j.1365-2893.2007.00871.x. — PMID 17927615. — PMC 2156112.
- ↑ Rudin D., Shah S. M., Kiss A., Wetz R. V., Sottile V. M. Interferon and lamivudine vs. interferon for hepatitis B e antigen-positive hepatitis B treatment: meta-analysis of randomized controlled trials (англ.) // Liver Int. : journal. — 2007. — Vol. 27, no. 9. — P. 1185—1193. — doi:10.1111/j.1478-3231.2007.01580.x. — PMID 17919229. — PMC 2156150.
- ↑ Dimmock, 2007, p. 3.
- ↑ Goris N., Vandenbussche F., De Clercq K. Potential of antiviral therapy and prophylaxis for controlling RNA viral infections of livestock (англ.) // Antiviral Res. : journal. — 2008. — Vol. 78, no. 1. — P. 170—178. — doi:10.1016/j.antiviral.2007.10.003. — PMID 18035428.
- ↑ Carmichael L. An annotated historical account of canine parvovirus (англ.) // J. Vet. Med. B Infect. Dis. Vet. Public Health : journal. — 2005. — Vol. 52, no. 7—8. — P. 303—311. — doi:10.1111/j.1439-0450.2005.00868.x. — PMID 16316389.
- ↑ Tinsley T. W., Harrap K. A. Viruses of Invertebrates (англ.). Дата обращения: 27 февраля 2013. Архивировано 7 июня 2018 года.
- ↑ Invertebrate iridescent viruses (Iridoviridae) (англ.). Дата обращения: 27 февраля 2013. Архивировано 13 сентября 2019 года.
- ↑ Вирусы беспозвоночных. Дата обращения: 27 февраля 2013. Архивировано 24 февраля 2020 года.
- ↑ Chen Y. P., Zhao Y., Hammond J., Hsu H., Evans J. D., Feldlaufer M. F. Multiple virus infections in the honey bee and genome divergence of honey bee viruses (англ.) // Journal of Invertebrate Pathology : journal. — Vol. 87, no. 2—3. — P. 84—93. — doi:10.1016/j.jip.2004.07.005. — PMID 15579317.
- ↑ Rybicki EP (2015) «A Top Ten list for economically important plant viruses», Archives of Virology 160:17-20
- ↑ Shors, 2008, p. 584.
- ↑ Shors, 2008, pp. 562—587.
- ↑ Dinesh-Kumar S. P., Tham Wai-Hong, Baker B. J. Structure—function analysis of the tobacco mosaic virus resistance gene N (англ.) // Proceedings of the National Academy of Sciences : journal. — National Academy of Sciences, 2000. — Vol. 97, no. 26. — P. 14789—14794. — doi:10.1073/pnas.97.26.14789. — PMID 11121079. — PMC 18997. Архивировано 19 марта 2017 года.
- ↑ Shors, 2008, pp. 573—576.
- ↑ Soosaar J. L., Burch-Smith T. M., Dinesh-Kumar S. P. Mechanisms of plant resistance to viruses // Nat. Rev. Microbiol. — 2005. — Т. 3, № 10. — С. 789—798. — doi:10.1038/nrmicro1239. — PMID 16132037.
- ↑ Lomonossoff G. P. Virus Particles and the Uses of Such Particles in Bio- and Nanotechnology // Recent Advances in Plant Virology. — Caister Academic Press (англ.) (рус., 2011. — ISBN 978-1-904455-75-2.
- ↑ Упадышев М. Т. Оздоровление садовых растений от вирусов // Защита и карантин растений. — 2012-01-01. — Вып. 5. — ISSN 1026-8634. (недоступная ссылка)
- ↑ 1 2 3 Вирусы грибов. Дата обращения: 27 февраля 2013. Архивировано 5 сентября 2019 года.
- ↑ McGraw-Hill Science & Technology Encyclopedia: Fungal virus (англ.). Дата обращения: 27 февраля 2013. Архивировано 4 марта 2016 года.
- ↑ Дьяков Ю. Т., Шнырева А. В., Сергеев А. Ю. Введение в генетику грибов. — М.: изд. центр «Академия», 2005. — С. 57—58. — 304 с. — ISBN 5-7695-2174-0.
- ↑ Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л. Ботаника: в 4 томах. — М.: изд. центр «Академия», 2006. — Т. 1. Водоросли и грибы. — С. 73. — 320 с. — ISBN 5-7695-2731-5.
- ↑ International Society for Viruses of Microorganisms: Protist Viruses (англ.). Дата обращения: 28 февраля 2013. (недоступная ссылка)
- ↑ Philippe Colson, Gregory Gimenez, Mickaël Boyer, Ghislain Fournous, Didier Raoult. The Giant Cafeteria roenbergensis Virus That Infects a Widespread Marine Phagocytic Protist Is a New Member of the Fourth Domain of Life (англ.). Архивировано 31 мая 2020 года.
- ↑ Wommack K. E., Colwell R. R. Virioplankton: viruses in aquatic ecosystems // Microbiol. Mol. Biol. Rev.. — 2000. — Т. 64, № 1. — С. 69—114. — doi:10.1128/MMBR.64.1.69-114.2000. — PMID 10704475. — PMC 98987.
- ↑ Bergh O., Børsheim K. Y., Bratbak G., Heldal M. High abundance of viruses found in aquatic environments (англ.) // Nature : journal. — 1989. — Vol. 340, no. 6233. — P. 467—468. — doi:10.1038/340467a0. — PMID 2755508.
- ↑ Shors, 2008, pp. 595—597.
- ↑ Bickle T. A., Krüger D. H. Biology of DNA restriction (англ.) // Microbiology and Molecular Biology Reviews (англ.) (рус.. — American Society for Microbiology (англ.) (рус., 1993. — 1 June (vol. 57, no. 2). — P. 434—450. — PMID 8336674. — PMC 372918.
- ↑ Barrangou R., Fremaux C., Deveau H. et al. CRISPR provides acquired resistance against viruses in prokaryotes (англ.) // Science. — 2007. — Vol. 315, no. 5819. — P. 1709—1712. — doi:10.1126/science.1138140. — PMID 17379808.
- ↑ Brouns S. J., Jore M. M., Lundgren M., et al. Small CRISPR RNAs guide antiviral defense in prokaryotes (англ.) // Science. — 2008. — Vol. 321, no. 5891. — P. 960—964. — doi:10.1126/science.1159689. — PMID 18703739.
- ↑ В. М. Жданов. Человек и вирусы // Наука и человечество, 1984. — М.: Знание. — С. 44—55.
- ↑ Xiang X., Chen L., Huang X., Luo Y., She Q., Huang L. Sulfolobus tengchongensis spindle-shaped virus STSV1: virus-host interactions and genomic features (англ.) // J. Virol. : journal. — 2005. — Vol. 79, no. 14. — P. 8677—8686. — doi:10.1128/JVI.79.14.8677-8686.2005. — PMID 15994761. — PMC 1168784. Архивировано 16 октября 2019 года.
- ↑ Robb F., Antranikian G., Grogan D., Driessen A. (Editors) (2007) «Thermophiles: Biology and Technology at High Temperatures», CRC Press, ISBN 978-0849392146, pp. 231—232
- ↑ Krupovic M., Quemin E. R. J., Bamford D. H., Forterre P., Prangishvili D. Unification of the Globally Distributed Spindle-Shaped Viruses of the Archaea // Journal of Virology. — 2013. — 11 декабря (т. 88, № 4). — С. 2354—2358. — ISSN 0022-538X. — doi:10.1128/JVI.02941-13. [исправить]
- ↑ Hochstein Rebecca, Bollschweiler Daniel, Engelhardt Harald, Lawrence C. Martin, Young Mark. Large Tailed Spindle Viruses of Archaea: a New Way of Doing Viral Business // Journal of Virology. — 2015. — 17 июня (т. 89, № 18). — С. 9146—9149. — ISSN 0022-538X. — doi:10.1128/JVI.00612-15. [исправить]
- ↑ Uneven Distribution of Viruses Suggests Surprising Evolutionary Power Архивная копия от 8 августа 2017 на Wayback Machine // Scientific American
- ↑ Prangishvili D., Forterre P., Garrett R. A. Viruses of the Archaea: a unifying view // Nat. Rev. Microbiol.. — 2006. — Т. 4, № 11. — С. 837—848. — doi:10.1038/nrmicro1527. — PMID 17041631.
- ↑ Prangishvili D., Garrett R.A. Exceptionally diverse morphotypes and genomes of crenarchaeal hyperthermophilic viruses (англ.) // Biochem. Soc. Trans. : journal. — 2004. — Vol. 32, no. Pt 2. — P. 204—208. — doi:10.1042/BST0320204. — PMID 15046572. (недоступная ссылка)
- ↑ Mojica F. J., Díez-Villaseñor C., García-Martínez J., Soria E. Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements (англ.) // J. Mol. Evol. : journal. — 2005. — Vol. 60, no. 2. — P. 174—182. — doi:10.1007/s00239-004-0046-3. — PMID 15791728.
- ↑ Makarova K. S., Grishin N. V., Shabalina S. A., Wolf Y. I., Koonin E. V. A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action (англ.) // Biol. Direct : journal. — 2006. — Vol. 1. — P. 7. — doi:10.1186/1745-6150-1-7. — PMID 16545108. — PMC 1462988. Архивировано 10 мая 2013 года.
- ↑ 1 2 La Scola B., Desnues C., Pagnier I., Robert C., Barrassi L., Fournous G., Merchat M., Suzan-Monti M., Forterre P., Koonin E., Raoult D. The virophage as a unique parasite of the giant mimivirus (англ.) // Nature. — 2008. — Vol. 455, iss. 7209. — P. 100—104. — doi:10.1038/nature07218. — PMID 18690211.
- ↑ Helen Pearson. ‘Virophage’ suggests viruses are alive (2008). Дата обращения: 2 февраля 2013. Архивировано 10 февраля 2013 года.
- ↑ Александр Марков. Вирусы тоже болеют вирусными заболеваниями (8 сентября 2008). Дата обращения: 2 февраля 2013. Архивировано 10 февраля 2013 года.
- ↑ Desnues C., Boyer M., Raoult D. Sputnik, a virophage infecting the viral domain of life // Adv Virus Res. — 2012. — Т. 82. — С. 63—89. — doi:10.1016/B978-0-12-394621-8.00013-3. — PMID 22420851.
- ↑ Claverie J. M., Abergel C. Mimivirus and its virophage // Annu Rev Genet. — 2009. — Т. 43. — С. 49—66. — doi:10.1146/annurev-genet-102108-134255. — PMID 19653859.
- ↑ Zhou Jinglie, Zhang Weijia, Yan Shuling, Xiao Jinzhou, Zhang Yuanyuan, Li Bailin, Pan Yingjie, Wang Yongjie. Diversity of Virophages in Metagenomic Data Sets // Journal of Virology. — 2013. — Vol. 87, no. 8. — P. 4225—4236. — doi:10.1128/JVI.03398-12. — PMID 23408616. [исправить]
- ↑ Zhang X., Sun S., Xiang Y., Wong J., Klose T., Raoult D., Rossmann M. G. Structure of Sputnik, a virophage, at 3.5-Å resolution. (англ.) // Proceedings of the National Academy of Sciences of the United States of America. — 2012. — Vol. 109, no. 45. — P. 18431—18436. — doi:10.1073/pnas.1211702109. — PMID 23091035. [исправить]
- ↑ La Scola B., Campocasso A., N’Dong R., Fournous G., Barrassi L., Flaudrops C., Raoult D. Tentative characterization of new environmental giant viruses by MALDI-TOF mass spectrometry // Intervirology. — 2010. — Т. 53, вып. 5. — С. 344—353. — doi:10.1159/000312919. — PMID 20551686.
- ↑ Bekliz M., Colson P., La Scola B. The Expanding Family of Virophages // Viruses. — 2016. — Vol. 8, no. 11. — doi:10.3390/v8110317. — PMID 27886075. [исправить]
- ↑ ScienceNow — «Ancient Virus Gave Wasps Their Sting». Дата обращения: 26 марта 2011. Архивировано из оригинала 15 апреля 2011 года.
- ↑ Элементы — новости науки: Наездники подавляют иммунную защиту своих жертв при помощи прирученных вирусов. Дата обращения: 19 августа 2009. Архивировано 17 ноября 2016 года.
- ↑ Breitbart M. Marine viruses: truth or dare // Annual Review of Marine Science. — 2012. — Т. 4. — С. 425—448. — PMID 22457982.
- ↑ Shors, 2008, p. 4.
- ↑ Shors, 2008, p. 5.
- ↑ Shors, 2008, p. 593.
- ↑ Suttle C. A. Viruses in the sea (англ.) // Nature. — 2005. — Vol. 437, no. 7057. — P. 356—361. — doi:10.1038/nature04160. — PMID 16163346. (недоступная ссылка)
- ↑ National Center for Environmental Health. Harmful Algal Blooms (англ.) (недоступная ссылка — история). Centers for disease control and prevention (24 июля 2012). Дата обращения: 8 марта 2013. Архивировано 21 марта 2013 года.
- ↑ 1 2 3 4 Suttle C. A. Marine viruses—major players in the global ecosystem (англ.) // Nature Reviews. Microbiology : journal. — 2007. — Vol. 5, no. 10. — P. 801—812. — doi:10.1038/nrmicro1750. — PMID 17853907. Архивировано 4 марта 2016 года.
- ↑ Hall, A. J., Jepson, P. D., Goodman, S. J. & Harkonen, T. «Phocine distemper virus in the North and European Seas — data and models, nature and nurture». Biol. Conserv. 131, 221—229 (2006).
- ↑ Forterre P., Philippe H. The last universal common ancestor (LUCA), simple or complex? (англ.) // The Biological Bulletin : journal. — 1999. — Vol. 196, no. 3. — P. 373—375. — doi:10.2307/1542973. — PMID 11536914.
- ↑ Collier, 1998, p. 8.
- ↑ Lodish, Harvey; Berk, Arnold; Zipursky, S. Lawrence; Matsudaira, Paul; Baltimore, David; Darnell, James. Viruses:Structure, Function, and Uses Retrieved on September 16, 2008
- ↑ Matsuzaki S., Rashel M., Uchiyama J., Sakurai S., Ujihara T., Kuroda M., Ikeuchi M., Tani T., Fujieda M., Wakiguchi H., Imai S. Bacteriophage therapy: a revitalized therapy against bacterial infectious diseases (англ.) // Journal of Infection and Chemotherapy : Official Journal of the Japan Society of Chemotherapy : journal. — 2005. — October (vol. 11, no. 5). — P. 211—219. — doi:10.1007/s10156-005-0408-9. — PMID 16258815.
- ↑ Gleba, Y. Y.; Giritch, A. Plant Viral Vectors for Protein Expression // Recent Advances in Plant Virology. — Caister Academic Press (англ.) (рус., 2011. — ISBN 978-1-904455-75-2.
- ↑ Fischlechner M., Donath E. Viruses as Building Blocks for Materials and Devices (англ.) // Angewandte Chemie International Edition : journal. — 2007. — Vol. 46, no. 18. — P. 3184—3193. — doi:10.1002/anie.200603445. — PMID 17348058.
- ↑ Soto C. M., Blum A. S., Vora G. J., et al. Fluorescent signal amplification of carbocyanine dyes using engineered viral nanoparticles (англ.) // J. Am. Chem. Soc. : journal. — 2006. — Vol. 128, no. 15. — P. 5184—5189. — doi:10.1021/ja058574x. — PMID 16608355.
- ↑ Blum A. S., Soto C. M., Wilson C. D. et al. An Engineered Virus as a Scaffold for Three-Dimensional Self-Assembly on the Nanoscale (англ.) // Small : journal. — 2005. — Vol. 7. — P. 702. — doi:10.1002/smll.200500021. — PMID 17193509.
- ↑ Coleman J. R., Papamichail D., Skiena S., Futcher B., Wimmer E., Mueller S. Virus attenuation by genome-scale changes in codon pair bias (англ.) // Science : journal. — 2008. — Vol. 320, no. 5884. — P. 1784—1787. — doi:10.1126/science.1155761. — PMID 18583614. — PMC 2754401.
- ↑ Genomes. NIH viral genome database. Ncbi.nlm.nih.gov. Дата обращения: 7 мая 2012. Архивировано 14 февраля 2014 года.
- ↑ Shors, 2008, p. 331.
- ↑ 1 2 3 Artenstein A. W., Grabenstein J. D. Smallpox vaccines for biodefense: need and feasibility (англ.) // Expert Review of Vaccines : journal. — 2008. — Vol. 7, no. 8. — P. 1225—1237. — doi:10.1586/14760584.7.8.1225. — PMID 18844596. Архивировано 2 июля 2016 года.
- ↑ Aragón T. J., Ulrich S., Fernyak S., Rutherford G. W. Risks of serious complications and death from smallpox vaccination: a systematic review of the United States experience, 1963–1968 (англ.) // BMC public health : journal. — 2003. — Vol. 3. — P. 26. — doi:10.1186/1471-2458-3-26. — PMID 12911836. — PMC 194634. Архивировано 4 октября 2013 года.
- ↑ Weiss M. M., Weiss P. D., Mathisen G., Guze P. Rethinking smallpox // Clin. Infect. Dis.. — 2004. — Т. 39, № 11. — С. 1668—1673. — doi:10.1086/425745. — PMID 15578369.
- ↑ 1 2 3 4 Georgios Pappas, Savvas Seitaridis, Nikolaos Akritidis, Epaminondas Tsianos. Infectious Diseases in Cinema: Virus Hunters and Killer Microbes (англ.) // Clinical Infectious Diseases : Oxford Journals. — 2003. — Vol. 37, no. 7. — P. 939—942. — doi:10.1086/377740. Архивировано 20 октября 2014 года.
Литература[править | править код]
- Букринская А. Г. Вирусология : Учеб. пособие. — М.: Медицина, 1986. — 336 с.
- Зуев В. А. Многоликий вирус. — М.: АСТ, 2020. — 304 с. — (Наука и жизнь). — 3000 экз. — ISBN 978-5-17-118736-1.
- Mayo M. A., Pringle C. R. Virus taxonomy — 1997 (англ.) // Journal of General Virology (англ.) (рус.. — Microbiology Society (англ.) (рус., 1998. — No. 79. — P. 649—657. Архивировано 29 сентября 2007 года.
- Collier, Leslie; Balows, Albert; Sussman, Max. Topley and Wilson’s Microbiology and Microbial Infections / Mahy, Brian and Collier, Leslie. Arnold. — ninth edition. — Virology, 1998. — Т. 1. — ISBN 0-340-66316-2.
- Dimmock N. J., Easton, Andrew J.; Leppard, Keith. Introduction to Modern Virology. — sixth edition. — Blackwell Publishing, 2007. — ISBN 1-4051-3645-6.
- Knipe, David M.; Howley, Peter M.; Griffin, Diane E.; Lamb, Robert A.; Martin, Malcolm A.; Roizman, Bernard; Straus Stephen E. Fields Virology. — Lippincott Williams & Wilkins., 2007. — ISBN 0-7817-6060-7.
- Shors, Teri. Understanding Viruses. — Jones and Bartlett Publishers, 2008. — ISBN 0-7637-2932-9.
- The International Code of Virus Classification and Nomenclature : October 2018 : [англ.] // ICTV. — 2018. (Дата обращения: 17 мая 2020).
Ссылки[править | править код]
- The Universal Virus Database ICTVdB. Дата обращения: 13 декабря 2007. Архивировано из оригинала 13 декабря 2007 года.
- ViralZone Ресурс Швейцарского института биоинформатики, предоставляющий информацию обо всех семействах вирусов, сопровождается общей молекулярной и эпидемиологической информацией. (англ.)
- Онлайн-семинар Дейвида Балтимора: «Введение в вирусы и ВИЧ». Дата обращения: 13 октября 2011. Архивировано из оригинала 13 октября 2011 года. (англ.)
- Онлайн-семинар Эри Гелениуса: «Введение в вирусы». Дата обращения: 14 октября 2011. Архивировано из оригинала 14 октября 2011 года. (англ.)
- 3D-структуры вирусов в EM Data Bank (EMDB). (англ.)
- Пташник, Ольга; Волкова, Ольга. Вирусы. Раскрась вирусные частицы. // Сайт Biomolecula.ru (17 февраля 2017). Дата обращения: 5 апреля 2018.
- Лапочкин, Юрий. Вирусные геномы в системе эволюции. // Сайт Biomolecula.ru (28 ноября 2014). Дата обращения: 5 апреля 2018.
- Ржешевский, Алексей. Вирусы и человек. Противостояние длиной в тысячелетия. // Сайт Biomolecula.ru (8 ноября 2015). Дата обращения: 5 апреля 2018.
- Чугунов, Антон. Нобелевскую премию 2008 года по физиологии и медицине вручили за вирусологические исследования. // Сайт Biomolecula.ru (6 ноября 2008). Дата обращения: 5 апреля 2018.
- Ожаровская, Татьяна. Фаговый домик. А в ваши строительные магазины уже завезли фаговые нанопровода? // Сайт Biomolecula.ru (11 ноября 2016). Дата обращения: 5 апреля 2018.
- Закубанский, Александр. Вирусы-платформы: яд во благо. // Сайт Biomolecula.ru (5 октября 2012). Дата обращения: 5 апреля 2018.
- Минина, Елизавета. Анти-CRISPR: ответ вирусов. // Сайт Biomolecula.ru (6 декабря 2017). Дата обращения: 5 апреля 2018.
- Цветные изображения вирусов из книги Карла Циммера Планета вирусов (2011) .
Как легко найти вирус на компьютере и избавиться от него
На чтение 4 мин Опубликовано 02.08.2020
Заражение вирусом – неприятная проблема, с которой сталкиваются все владельцы ПК, мобильных устройств. Многообразие вредоносных программ поражает. Есть вирусы, которые просто мешают работать, «притормаживают» компьютер, а есть и более сложные программы, например, ворующие личные данные или передающие их в сеть. Обнаружить и удалить вирус сложно, но можно. Пригодятся определенные знания и немного терпения. Также нелишним будет усвоить правила безопасности – профилактика всегда лучше «лечения».

Содержание
- Как обнаружить вирусы на устройстве
- Поиск и установка антивирусных программ
- Лучшие программы для обнаружения вируса
- Что делать если вирус не удаляется
- Меры профилактики от заражения вирусом
Как обнаружить вирусы на устройстве
Чтобы стать профессионалом в области компьютерных программ, придется долго учиться. Но для поиска и удаления простейших вирусов знания ВУЗа не понадобятся, с работой справятся антивирусные программы. Их бывает много, все они разные, но цель одна – выявить, обезвредить и уничтожить вирус до того, как он станет причиной разрушения файлов.
Поиск и установка антивирусных программ
Простейшие антивирусники бесплатно доступны в сети. Чтобы найти программу, в строку браузера забивают «антивирусная программа для ПК (ноутбука, планшета)» и выбирают из списка ПО, наиболее подходящее по параметрам. Установка не отнимет много времени, проходит автоматически:
- нажимают на кнопку Скачать;
- указывают адрес инсталляции софта (диск D, C);
- запускают скачивание;
- следуют инструкции по установке.
После установки программы компьютер проверяют на наличие вирусов. Если ПО обнаружит нарушения, отобразит его в окошке. Пользователю нужно выбрать действие: устранить/игнорировать.
Профессиональные лицензионные антивирусные программы всегда платные. Преимущества таких ПО многочисленны:
- Программа регулярно обновляется в автоматическом режиме. Это значит, что владелец устройства получает более качественную защиту.
- Проверка на вирусы проводится в автоматическом режиме.
- При обнаружении вирусов, софт выполнит заданные действия – удалит, переместит в карантин и т.д.

Вот так выглядит антивирусная программа в действии
Лучшие программы для обнаружения вируса
Специалисты не дают рекомендаций относительно лучших или худших программ – все относительно, выбор строго индивидуальный. Специалисты настоятельно рекомендуют скачать на устройство сканер вирусов, например, Malwarebytes. Программа занимает немного места и хорошо отслеживает различные нарушения.
Сканер не панацея, он может пропустить какие-то особо въедливые программы – руткиты. Поиск этих вредоносных ПО поручают Sophos Virus Removal Tool или Malwarebytes Anti-Rootkit. Эти программы умеют обнаруживать самые сложные вирусы. Также пользователю пригодится AVG Rescue CD – сканер, который запускают перед началом проверки на вирусы. Программа предотвращает последствия сбоев работы из-за вирусов, помогает восстановить поломанные, поврежденные, утерянные файлы.

Что делать если вирус не удаляется
Вариантов решения проблемы два:
- Переустановить операционную систему. Перед этим данные с устройства копируют переносом на другие носители. Затем запускают очистку, переустановку. Процедура требует опыта, знаний, поэтому обычный пользователь может и не справиться.
- Отнести устройство в сервис. Специалисты оценят возможность восстановления файлов, очистки компьютера без переустановки. Но если мастер скажет, что нужно переустановить, надо соглашаться.
Многие владельцы ноутбуков не обращаются к мастерам по причине высокой стоимости услуги. Однако тут стоит взвесить все особенности, самостоятельная переустановка с последующим восстановлением файлов может обойтись намного дороже.
Меры профилактики от заражения вирусом
Чтобы не допустить заражения, специалисты советуют придерживаться нескольких простых правил:
- Не переходить по сомнительным ссылкам. Такие сайты всегда заражены вирусами.
- Обязательно скачать и установить антивирусную программу. Лучше всего отдать предпочтение проверенным лицензионным ПО с регулярным обновлением.
- Не сохранять на сайтах пароли, номера карт банка и прочие личные данные. Именно за ними охотятся хакеры, данные нужны для различных махинаций.
- Периодически менять пароли на всех сайтах, чтобы затруднить доступ к личным данным.
Главный совет – не пренебрегать помощью специалистов. Вмешательство в работу ноутбука или планшета неопытного мастера может привести к окончательной поломке гаджета.

Оцените качество статьи. Нам важно ваше мнение:
![]()
: 29 сен 2014 , Мой НГУ , том 57/58,
№3/4
Вирус под микроскопом: от визуализации к манипуляции
Вирусы являются чрезвычайно малыми объектами, имеющими размер от нескольких десятков до нескольких сотен нанометров. Первым и на несколько десятилетий единственным методом их визуализации стала электронная микроскопия, позволившая не только подробно изучить строение самих вирусных частиц, называемых вирионами, но и исследовать процессы, происходящие в зараженной клетке – репликацию вируса. «Монополия» электронной микроскопии была нарушена появлением в начале 1980-х годов принципиально нового класса приборов – сканирующих зондовых микроскопов.
Относящийся к данному классу атомно-силовой микроскоп оказался инструментом, подходящим для исследования биологических объектов и позволил не только визуализировать наноразмерные структуры, но и манипулировать ими. В частности, принципиально возможной оказалась манипуляция одиночными вирионами и прямое измерение сил, возникающих при их контакте с поверхностью клетки. Такие эксперименты позволяют получать подробные данные о самом первом и во многих случаях еще недостаточно исследованном этапе заражения клетки – адгезии вируса к ее поверхности. Данные исследования представляют и значительный практический интерес, т.к. могут дать ключ к созданию эффективных противовирусных препаратов, защищающих клетки от проникновения вирусов.
В известной песне Владимира Высоцкого поется: «не поймаешь нейтрино за бороду и не посадишь в пробирку…». Конечно, вирус – это не нейтрино, не атом и даже не молекула, но все же объект настолько малый, что его невозможно увидеть не только глазом, но и в обычный световой микроскоп. Однако электронная микроскопия, в сотни тысяч раз увеличившая возможности нашего зрения, позволила не только увидеть эти удивительные объекты, но и рассмотреть их до мельчайших подробностей. А атомно-силовая микроскопия, в такой же степени обострившая наше осязание, позволила осуществить прямую механическую манипуляцию вирусными частицами
Вирусы являются чрезвычайно малыми объектами – их размеры лежат в диапазоне от нескольких десятков до нескольких сотен нанометров. Первым и на долгое время единственным методом прямой визуализации наноразмерных частиц стала электронная микроскопия (ЭМ), которая начала развиваться в 1930-е гг. Метод, оказавшийся очень информативным, позволил не только детально охарактеризовать структуру различных вирусов, но и исследовать процессы, происходящие в зараженной клетке.
Оказалось, что форма вирусных частиц отличается большим разнообразием: от правильных сфер до сложных структур, напоминающих кирпичи, обклеенные трубочками (вирус натуральной оспы), или щетинистых червей (вирус геморрагической лихорадки Эбола).
Еще большее разнообразие было обнаружено для стратегии репликации (размножения) вирусов. Единственным фундаментальным свойством, общим для всех без исключения вирусов, оказался их статус облигатного внутриклеточного паразита. Это означает, что для размножения вируса его генетический материал должен в обязательном порядке проникнуть в живую клетку и «захватить» ее ферментативный аппарат, переключив последний на производство копий вируса.
Вне клетки любой вирус является всего лишь молекулярным контейнером с генетическим материалом (ДНК или РНК) и вряд ли может считаться полноценным живым организмом, хотя по этому вопросу в научной среде до сих пор нет окончательной терминологической определенности.
Спецификой электронной микроскопии является изучение фиксированных, т. е. подготовленных специальным образом, объектов – по сути, она работает только с «мертвой» материей *. Имея дело только с «застывшими мгновениями», исследователь может лишь строить гипотезы о динамике изучаемых процессов, поскольку не имеет возможности наблюдать их течение в реальном времени.
Так, исследование репликации вируса методом просвечивающей электронной микроскопии на ультратонких срезах выглядит следующим образом: зараженные клетки обрабатывают фиксирующим раствором, обезвоживают спиртом и заливают специальной смолой. После отвердевания смолы с помощью специального прибора – ультратома – делают ультратонкие (≈ 50 нм) срезы, которые затем наносят на специальную сетку и обрабатывают растворами солей тяжелых металлов. Во время самого микроскопического исследования образец находится в вакуумной камере и подвергается действию пучка электронов с энергией в несколько десятков кэВ. Очевидно, что прижизненная визуализация в данном случае принципиально невозможна.
В течение почти полувека электронная микроскопия оставалась единственным методом визуализации наноразмерных объектов. Однако в начале 1980-х гг. эта монополия была нарушена появлением сканирующей зондовой микроскопии (СЗМ). Основным принципом СЗМ является сканирование – прецизионное (с высокой точностью) перемещение зонда вблизи исследуемой поверхности, сопряженное с отслеживанием определенного параметра, характеризующего взаимодействие между зондом и образцом. Результатом такого сканирования является топографическая карта рельефа поверхности образца.
Первым прибором СЗМ стал сканирующий туннельный микроскоп (СТМ), который мог лишь весьма ограниченно использоваться для визуализации биологических объектов, так как для его работы требовалась высокая электрическая проводимость исследуемой поверхности.
В 1986 г. швейцарский физик Г. Бинниг и его коллеги создали новый прибор семейства СЗМ – атомно-силовой микроскоп (АСМ). В основе его работы лежит силовое (Ван-дер-Ваальсово) взаимодействие атомов зонда и поверхности. АСМ не требуется электрическая проводимость поверхности образца, и он может осуществлять съемку в жидкой среде. Поэтому этот прибор оказался удобным инструментом для исследования биологических объектов.
. Чувствительным элементом АСМ является упругая консоль (кантилевер), на конце которой закреплен острый зонд. Силы, возникающие между атомами острия зонда и исследуемой поверхностью приводят к деформации кантилевера, которая в свою очередь фиксируется при помощи оптической системы, реализованной в большинстве современных АСМ, на основе полупроводникового лазера и четырехсекционного фотоприемника. Размер кантилевера – 100÷300 × 20÷40 мкм при толщине около 2 мкм. Высота зонда – около 10 мкм")
С момента появления атомно-силового микроскопа было опубликовано огромное число работ, посвященных АСМ-визуализации самых разнообразных биологических образцов. Следует все же признать, что в большинстве случаев в плане визуализации АСМ не дает ничего принципиально нового в сравнении с обычной электронной микроскопией, поэтому зачастую данный метод воспринимается биологами как техническая экзотика, а не как полноценный исследовательский инструмент.
Однако важнейшим, пусть и почти единственным преимуществом визуализации биологических объектов при помощи АСМ по сравнению с электронной микроскопией является возможность выполнения исследований нативных, природных образцов без какой-либо фиксации и специальной пробоподготовки, при физиологических параметрах среды.
Помимо визуализации рельефа поверхности с субнанометровым разрешением АСМ позволяет осуществлять прямое измерение сил, возникающих при взаимо¬действии одиночных наноразмерных объектов.
Проводятся такие измерения следующим образом: один объект закрепляется на острие зонда АСМ, а второй фиксируется на подложке, после чего зонд подводится к поверхности подложки до достижения механического контакта, а затем возвращается обратно. В ходе этого перемещения отслеживается деформация упругой консоли (кантилевера). Зависимость этого параметра от расстояния между зондом и подложкой называется силовой кривой. С ее помощью можно определить величину силы, действующей между исследуемыми объектами. Этот метод, названный атомно-силовой спектроскопией (АСС), может использоваться для исследования силовых характеристик взаимодействия самых разнообразных малых объектов: от неорганических наночастиц до вирусов и живых клеток.

Начальным этапом заражения клетки вирусом является адгезия (прилипание) вирусной частицы (вириона) к клеточной поверхности с последующим проникновением генетического материала вируса внутрь клетки. Этот процесс, определяемый взаимодействием белковых рецепторов, расположенных на поверхности клетки, с поверхностными белками вириона, является критически важным для размножения вируса. И, надо отметить, в большинстве случаев изучен недостаточно.
Поистине захватывающие перспективы исследований в этом направлении открывает АСС. Зафиксировав одиночную вирусную частицу на острие зонда АСМ, можно осуществить измерение силы, возникающей при контакте вирусной частицы с поверхностью клетки, исследовать кинетические характеристики данного взаимодействия и даже «вдавить» вирион внутрь клетки, одновременно ведя наблюдение при помощи мощного светового микроскопа. В таком эксперименте исследователь из пассивного наблюдателя превращается в активного участника процесса, осуществляя механическую манипуляцию исследуемым наноразмерным объектом – такую возможность не может предоставить ни один из других видов микроскопии.
 при создании площадки для посадки вириона, является электронная микроскопия, как сканирующая, так и просвечивающая: б – площадка на острие зонда для посадки крупной вирусной частицы; в – вирусоподобная частица, закрепленная на острие зонда. Просвечивающая электронная микроскопия (JEM 1400, Jeol, Япония)")
Однако фиксация одиночной вирусной частицы на острие зонда атомно-силового микроскопа является весьма непростой задачей. Для успешного проведения эксперимента требуется большая подготовительная работа:
- получить как можно более чистый и концентрированный препарат вируса;
- подготовить на острие зонда площадку подходящего размера для посадки вириона;
- химически активировать поверхность зонда для образования ковалентных связей при контакте с белками вируса;
- убедиться в том, что на зонде закрепился действительно вирион, а не молекулы свободного белка или мелкие фрагменты клеток, всегда присутствующие в препаратах вирусов.
Оценка концентрации и степени чистоты препарата вируса обычно проводится методом просвечивающей электронной микроскопии. Площадку на острие АСМ-зонда, которое обычно изготавливают из кремния или его нитрида, формируют путем длительного сканирования кремниевой или сапфировой подложки при больших значениях развертки и силы прижатия зонда к поверхности. Наиболее наглядной иллюстрацией для этого процесса служит изменение формы острия карандаша в ходе интенсивного рисования.
Главный вопрос, на который необходимо ответить при интерпретации любых результатов атомно-силовой спектроскопии, можно сформулировать следующим образом: «Силы между какими объектами были измерены?»
По меркам микроскопии, клетка высших организмов является относительно крупным (≈ 10 мкм) объектом, поэтому хорошо видна в световом микроскопе, при помощи которого на нее наводится кантилевер атомно-силового микроскопа. Но как быть с самим зондом, на острие которого предполагается наличие вириона? Строго говоря, вместо вириона там может оказаться все, что угодно: монослой белковых молекул, фрагмент клетки или вириона, агрегат из нескольких вирионов, случайное загрязнение и т. д. Кроме того, в процессе измерения вирион может разрушиться или оторваться от зонда. Визуализация же зонда с вирусной частицей методом электронной микроскопии до силовых измерений недопустима, так как под воздействием высушивания, вакуума и пучка электронов вирион приобретет необратимые изменения.
Наиболее эффективным методом решения данной проблемы оказалась визуализация острия зонда АСМ с помощью электронной микроскопии, осуществляемая непосредственно после силовых измерений. Если на острие будет обнаружена вирусная частица, уцелевшая в ходе эксперимента, то все сомнения развеются.
В течение последних пятидесяти лет в результате поистине титанической работы, проделанной электронными микроскопистами всего мира, накоплен огромный багаж знаний в области ультраструктурных аспектов репликации различных вирусов. Создание атомно-силового микроскопа и техники силовой спектроскопии позволило вплотную приблизиться к произвольной механической манипуляции одиночными вирусными частицами. Это выводит изучение взаимодействия вируса с клеткой на принципиально другой уровень – от структурных исследований к функциональным.
При этом атомно-силовая спектроскопия не является конкурентом для электронной микроскопии, а открывает новое самостоятельное направление исследований – наномеханику взаимодействия вирусной частицы с поверхностью клетки. Весьма вероятно, что в самом ближайшем будущем в данном направлении будут совершены фундаментальные открытия, соизмеримые по значимости с достижениями электронной микроскопии в середине прошлого века.
Изучение механизмов связывания вирусных частиц с поверхностью клетки вызывает значительный интерес не только с позиции фундаментальной науки, но и в контексте практических приложений. Более детальное понимание этих механизмов на молекулярном уровне может дать человечеству ключ к созданию эффективных противовирусных препаратов, защищающих клетки от проникновения вирусов.
*Просвечивающая электронная микроскопия с использованием специальной жидкостной ячейки и сканирующая электронная микроскопия при атмосферном давлении позволяют исследовать биологические объекты без фиксации, но из-за ряда технических трудностей и относительно низкого пространственного разрешения эти методы не получили широкого распространения
Литература
Корнеев Д. В., Бессуднова Е. В., Зайцев Б. Н. Изучение взаимодействия наночастиц TiO2 и поверхности эритроцитов человека методом атомно-силовой спектроскопии // УНЖ. 2012. № 4. С. 73—77.
Миронов В. Л. Основы сканирующей зондовой микроскопии. Нижний Новгород: ИФМ РАН, 2004. 182 с.
Alsteens D., Pesavent E., Cheuvart G. et al. Controlled manipulation of bacteriophages using single-virus force spectroscopy // ACSNANO. 2009. V. 3(10). P. 3063—3068.
Alsteens D., Trabelsi H., Soumillion P., Dufrene Y. F., Multiparametric atomic force microscopy imaging of single bacteriophages extruding from living bacteria // Nature Communications. V. 4. Article number: 2926.
Binnig G., Quate C. F., Gerber Ch. Atomic force microscope // Phys. Rev. Lett. 1986. V. 56(9). P. 930—933.
Cappella B., Dietler G. Force-distance curves by atomic force microscopy // Surf. Sci. Rep. 1999. V. 34. P. 1—104.
Malkin A.J., Plomp M., McPherson A. Unraveling the architecture of viruses by high-resolution atomic force microscopy // Methods Mol. Biol. 2005. V. 292. P. 85—108.
В публикации использованы фото автора
![]()
: 29 сен 2014 , Мой НГУ , том 57/58,
№3/4
Главная задача биологии — это развитие представлений у человека о живых организмах, о многообразии видов, обо всех закономерностях развития живых существ, а также об их взаимодействии с окружающей природой. Предмет основы безопасности жизнедеятельности (ОБЖ) позволяет получить знания и умения, которые помогут сохранить жизнь и здоровье в опасных ситуациях. Эти ситуации всегда возникают неожиданно, но, тем не менее, большинство из них предсказуемы и к ним можно подготовиться заранее. ОБЖ учит нас предвидеть возможные опасности и минимизировать потери от той или иной ситуации. Сегодня мы сталкиваемся с новым видом вирусной опасности COVID-19,о котором поговорим с точки зрения биологии и ОБЖ.
Что такое вирус?
Вирус — это неклеточный инфекционный агент. Сегодня нам известно около 6 тысяч различных вирусов, но их существует несколько миллионов. Вирусы не похожи друг на друга и могут иметь как форму сферы, спирали, так и форму сложного асимметричного сплетения. Размеры вирусов варьируются от 20 нм до 300 нм.
Как устроен вирус?
В центре агента находится генетический материал РНК или ДНК, вокруг которого располагается белковая структура — капсид.
Капсид служит для защиты вируса и помогает при захвате клетки. Некоторые вирусы дополнительно покрыты липидной оболочкой, т.е. жировой структурой, которая защищает их от изменений окружающей среды.
Вирусолог Дэвид Балтимор объединил все вирусы в 8 групп, из которых некоторые группы вирусов содержат 1-2 цепочки ДНК. Другие же содержат 1 цепочку РНК, которая может удваиваться или достраивать на своей матрице ДНК. При этом каждая группа вирусов производит себя в различных органеллах зараженной клетки.
Вирусы имеют определенный диапазон хозяев, т.е. он может быть опасен для одних видов и абсолютно безвреден для других. Например, оспой болеет только человек, а чумкой только некоторые виды плотоядных. Вирус не способен выжить сам по себе, поэтому активируется только в хозяйской клетке, используя ее ресурсы и питательные вещества. Цель вируса — создание множества копий себя, чтобы инфицировать другие клетки!

Вирусы. Цикл развития бактериофага. Скачать наглядное пособие в большом разрешении можно здесь.
Как вирус попадает в организм?
Вирусная инфекция начинается тогда, когда он проникает внутрь хозяина, а именно:
- через физические повреждения (например, порезы на коже)
- путём направленного впрыскивания (к примеру, укус комара)
- направленного поражения отдельной поверхности (например, при вдыхании вируса через трахею)
В зависимости от вида вируса, он может быть прикреплен:
- к эпителию слизистых оболочек (это например вирус гриппа)
- к нервной ткани (вирус простого герпеса)
- к иммунным клеткам (вирус иммунодефицита человека)

Биология. Рабочая тетрадь. 9 класс
Тетрадь является приложением к учебнику В. В. Пасечника, А. А. Каменского, Е. А. Криксунова, Г. Г. Швецова «Биология. Введение в общую биологию. 9 класс». Учебник соответствует ФГОС основного общего образования. Задания в тетради соответствуют содержанию разделов учебника и предназначены для самостоятельных работ учащихся с целью лучшего усвоения, систематизации и закрепления знаний, полученных при чтении учебника. В тетрадь включены тестовые задания, которые помогут ученикам подготовиться к успешной сдаче ЕГЭ и ГИА.
Купить
Геном вируса встраивается в одну из органелл или цитоплазму и превращает клетку в настоящий вирусный завод. Естественные процессы в клетке нарушаются, и она начинает заниматься производством и сбором белка вируса. Этот процесс называется репликацией. И его основная цель — это захват территории. Во время репликации генетический материал вируса смешивается с генами клетки хозяина — это приводит к активной мутации самого вируса, а также повышает его выживаемость. Когда процесс репликации налажен, вирусная частица отпочковывается и заражает уже новые клетки, в то время как инфицированная ранее клетка продолжает производство.
«Для проникновения в клетку белки поверхности вируса связываются со специфическими поверхностными белками клетки. Прикрепление, или адсорбция, происходит между вирусной частицей и клеточной мембраной. В мембране образуется дырка, и вирусная частица или только генетический материал попадают внутрь клетки, где будет происходить размножение вируса. Сегодня ученые всего мира сделали важное открытие о том, что заражение коронавирусом людей преклонного возраста объясняется тем, что у пожилых людей накапливается специфический белок, который помогает COVID-19 проникать внутрь клетки эпителия».
Кондратьева Елена
Выход вируса
Вирус создал множество собственных копий, клетка оказывается изнуренной из-за использования ее ресурсов. Больше вирусу клетка не нужна, поэтому клетка часто погибает и новорожденным вирусам приходится искать нового хозяина. Это и есть заключительная стадию жизненного цикла вируса.
Скорость распространения вирусной инфекции
Размножение вирусов протекает с исключительно высокой скоростью: при попадании в верхние дыхательные пути одной вирусной частицы уже через 8 часов количество инфекционного потомства достигает 10³, а концу первых суток − 10²³.
Вирусная латентность
Некоторые вирусы могут «спрятаться» внутри клетки. Это может происходить от того, чтобы уклониться от защитных реакций и иммунной системы хозяина, или просто от того, что продолжение репликации не входит в интересы вируса. Это умение прятаться называется латентностью. В течение определенного времени вирус не даёт начала потомкам и остается неактивным до тех пор, пока внешний стимул — например, свет или стресс, не активирует его.
Как вирус распространяется?
Существуют разные пути распространения вирусной инфекции.
- воздушно-капельный (кашель, чихание)
- с кожи на кожу (при прикосновениях и рукопожатиях)
- с кожи на продукты (при прикосновениях к пище грязными руками вирусы могут попасть в пищеварительную и дыхательную системы)
- через жидкие среды организма (кровь, слюну и другие)
Почему с вирусами так тяжело бороться?
Эволюция вирусов происходит буквально на наших глазах. Идет постоянная гонка между вирусами и живыми организмами. Эпидемии сопровождали человека с древних времён. Миллионы людей на различных континентах погибли от оспы и «испанского гриппа». Эпидемии этих болезней иногда были настолько опустошительными, что в некоторых городах, сёлах и деревнях умирало почти всё население. А когда вирус изобретает новое оружие — возникает пандемия.
Сегодня людям уже удалось победить некоторые вирусы, а некоторые взять под жесткий контроль. Например, Оспа (она же черная оспа). Болезнь вызывается вирусом натуральной оспы, передается от человека к человеку воздушно-капельным путем. Больные покрываются сыпью, переходящей в язвы, как на коже, так и на слизистых внутренних органов. Смертность, в зависимости от штамма вируса, составляет от 10 до 40 (иногда даже 70%), На сегодняшний день вирус полностью истреблен человечеством.
Кроме того, взяты под контроль такие заболевания, как бешенство, корь и полиомиелит. Но помимо этих вирусов существует масса других, которые требуют разработок или открытия новых вакцин.
Коронавирус
Виновником эпидемии, распространяющейся сегодня по миру, стал коронавирус, вирусная частица в 0,1 микрона. Свое название он получил благодаря наростам на своей структуре, своеобразным шипам. Внутри вируса спрятан яд, с помощью которого он подчиняет себе зараженный организм. Этот вирус воздействует не только на человека, но и на птиц, свиней, собак и летучих мышей. В настоящий момент выделяют от 30 до 39 разновидностей коронавирусной инфекции. Но для человека патогенно всего 6. И как любой другой вирус COVID-19 мутирует.

К наиболее распространенным симптомам COVID-19 относятся повышение температуры тела, сухой кашель и утомляемость. К более редким симптомам относятся боли в суставах и мышцах, заложенность носа, головная боль, конъюнктивит, боль в горле, диарея, потеря вкусовых ощущений или обоняния, сыпь и изменение цвета кожи на пальцах рук и ног. Как правило, эти симптомы развиваются постепенно и носят слабо выраженный характер. У некоторых инфицированных лиц болезнь сопровождается очень легкими симптомами.
Данные ВОЗ
Сколько же может жить этот вирус вне организма? Все зависит от типа вируса и от той поверхности, на которую вирусы попали. В качестве примера было рассмотрено 3 вируса, по которым велись исследования. Изучали время, на которое может задерживаться вирус на различных поверхностях. Данные приведены в таблице.

Поскольку пока не изобретено вакцины от COVID-19, в целях защиты от инфекции самым важным для нас является соблюдение гигиены.
Гигиена — раздел медицины, изучающий влияние жизни и труда на здоровье человека и разрабатывающая меры (санитарные нормы и правила), направленные на предупреждение заболеваний, обеспечение оптимальных условий существования, укрепление здоровья и продление жизни.
Сегодня следует соблюдать определенные правила гигиены:
- Соблюдение режима труда и отдыха, не допускающего развития утомления и переутомления.
- Выполнение условий, обеспечивающих здоровый и полноценный сон (свежий воздух, отсутствие шума, удобная постель, оптимальная продолжительность).
- Правильное здоровое питание в соответствии с потребностями организма.
- Комфортный микроклимат в жилище (температура, влажность и подвижность воздуха, естественная и искусственная освещенность помещений).
- Содержание в чистоте тела и тщательный уход за зубами.
- Спокойное и корректное поведение в конфликтных ситуациях.
Кроме вакцинации не стоит забывать о важных мерах предупреждения инфекционных болезней, таких как обеспечение безопасности воды, продовольственного сырья, продуктов питания, выполнение установленных санитарных требований в местах хранения и приготовления пищи, а также о строгом соблюдении правил личной гигиены (мытье рук, ношении масок и одноразовых перчаток на улице).

«Мы пытались рассказать Вам не только о существующих научных фактах о вирусах, но и показать, что определенные знания помогают нам в нынешней практической ситуации сохранить свое здоровье и здоровье своих близких. Мы понимаем, что сегодня коронавирус может находиться практически везде: на поверхностях любых предметов, в окружающей среде и т.д. Поэтому самоизоляция – это один из важнейших способов защиты от инфекции. Находясь дома, вы защищаете не только свое здоровье, но и помогаете медикам и ученым, которые сражаются с этим вирусов и день и ночь. Ведь, чем меньше шансов у нас с вами заболеть, тем больше шансов появляется у них, чтобы победить коронавирус. Пожалуйста, оставайтесь дома и соблюдайте режим самоизоляции и нормы гигиены».
Кондратьева Елена
