Эта тема входит в бесплатную часть моего курса по биохимии.
Белки — это полимерные молекулы, которые состоят из молекул поменьше — мономеров. Этими мономерами будут аминокислоты. Если упростить, то белок — это большой кирпичный дом. Где кирпичики — это аминокислоты. В этой статье мы посмотрим на эти кирпичики: какие они бывают, сколько их и какие у них свойства. Потом соединим аминокислоты вместе — синтезируем пептид. Подробно поговорим о пептидной связи, удерживающей аминокислоты вместе. А в конце небольшой подарок — торсионные углы.
Строение аминокислот
По названию понятно, что в аминокислоте должны быть две вещи: аминогруппа и карбоксильная группа. Аминокислот довольно много, но в белках встречаются только такие:
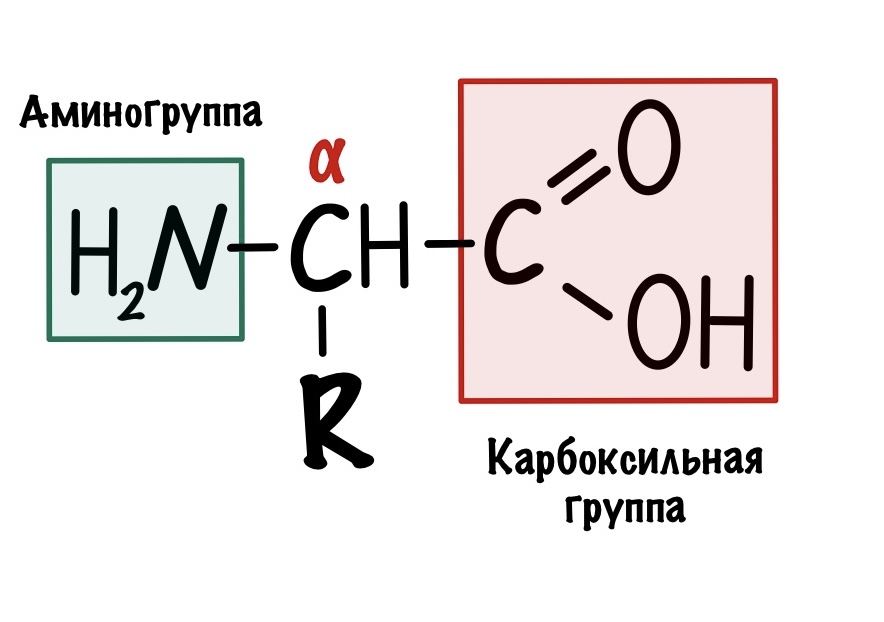
Это альфа-аминокислоты. В них есть центральный атом углерода, который связан с четырьмя заместителями: водородом, аминогруппой, карбоксильной группой и радикалом. Над этим атомом стоит греческая буква — альфа, почему? Это способ нумерации атомов углерода, входящих в органическое соединение. Для нумерации используют буквы греческого алфавита: альфа, бета, гамма, дельта и так далее. Нумерацию начинают с атома углерода, который следует после карбоксильной группы.
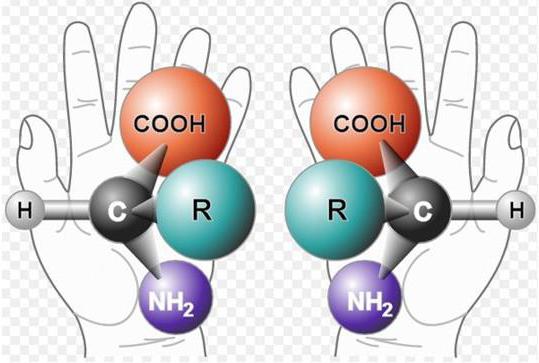
Альфа-углерод связан с четырьмя разными заместителями. Ещё разок повторим, что это — водород, аминогруппа, карбоксильная группа и радикал. Такие атомы называются хиральными центрами. Если в молекуле есть хиральный центр, то она хиральна. Что это значит? Это значит, что у молекулы есть изомер — он является её зеркальным отражением. Две этих молекулы будут несопоставимы в пространстве — они разные! Самый простой пример хиральности — левая и правая рука. Руки являются зеркальным отражением друг друга, но у нас не получится идеально сопоставить их в пространстве. Для этого пришлось бы превратить правую руку в левую. Или левую в правую.
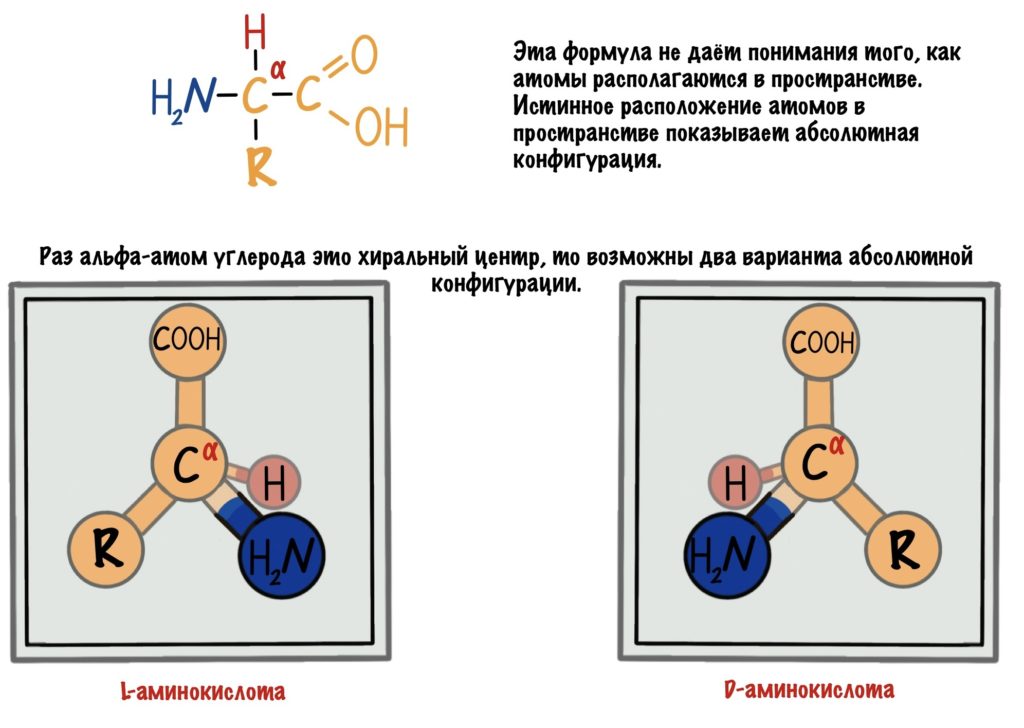
Живые организмы используют и синтезируют только L-аминокислоты.
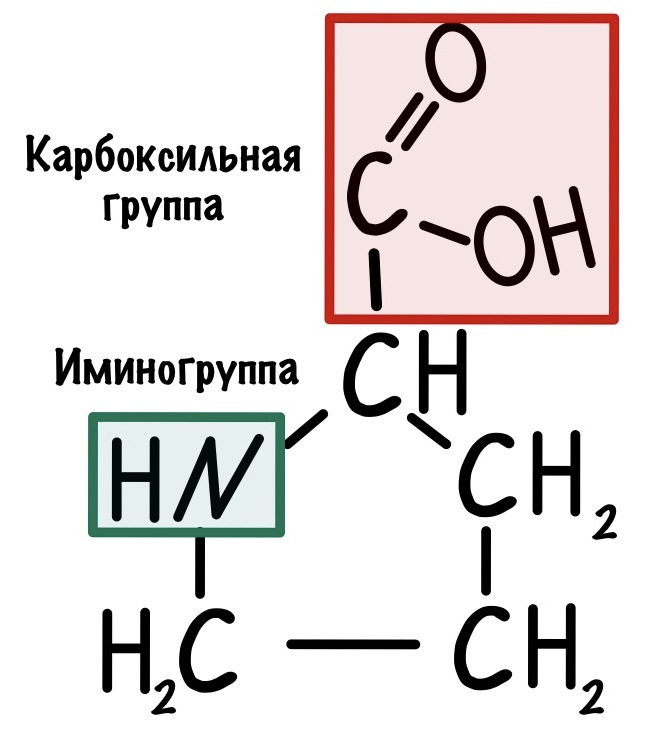
Альфа-аминокислоты будут различаться по своим радикалам. Всего нам нужно запомнить 20 аминокислот. Одна из них будет иметь особое строение — правильнее было бы назвать её иминокислотой. Вот наша легенда — пролин.
Вернемся к различиям между аминокислотами. Есть несколько классификаций радикалов, но мы возьмем самую полезную для нас — по полярности. А если говорить простыми словами, то по растворимости радикала в воде. Радикалы делятся на неполярные и полярные. Первые не растворимы в воде, а вторые растворимы. Когда будем говорить о строении белка, то поймем почему нас интересует именно эта классификация.
Неполярные радикалы аминокислот
У этих ребят нет групп, которые могут образовать водородные связи с водой, поэтому они нерастворимы. Вместо этого у них есть алифатические и ароматические группы. Радикалы выделены фиолетовым цветом.
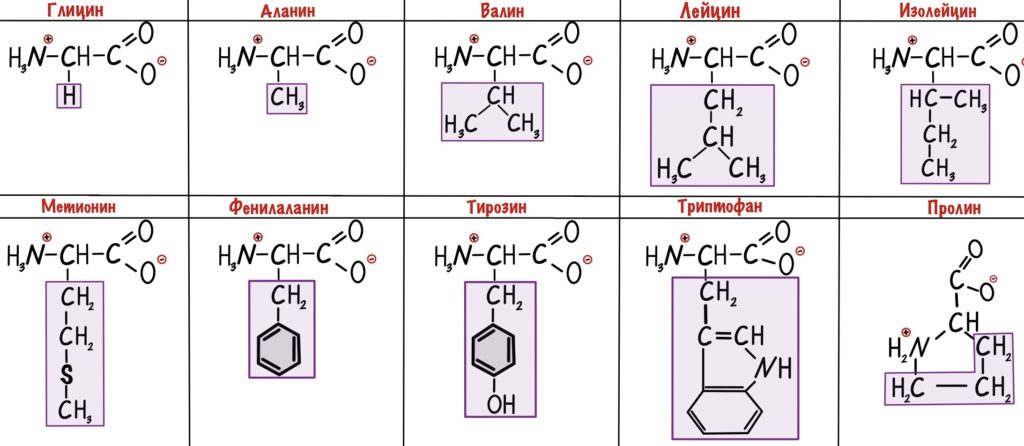
У глицина радикал — атом водорода. Выходит, что у его альфа-атома углерода два одинаковых заместителя — он не является хиральным атомом.
Полярные радикалы аминокислот
Перед этим остановимся на одной вещичке. Я писал формулы аминокислот так, как будто они не находятся в растворе. Но если мы заглянем в клетку, pH в цитоплазме которой 7 и 0, то увидим такую картину.
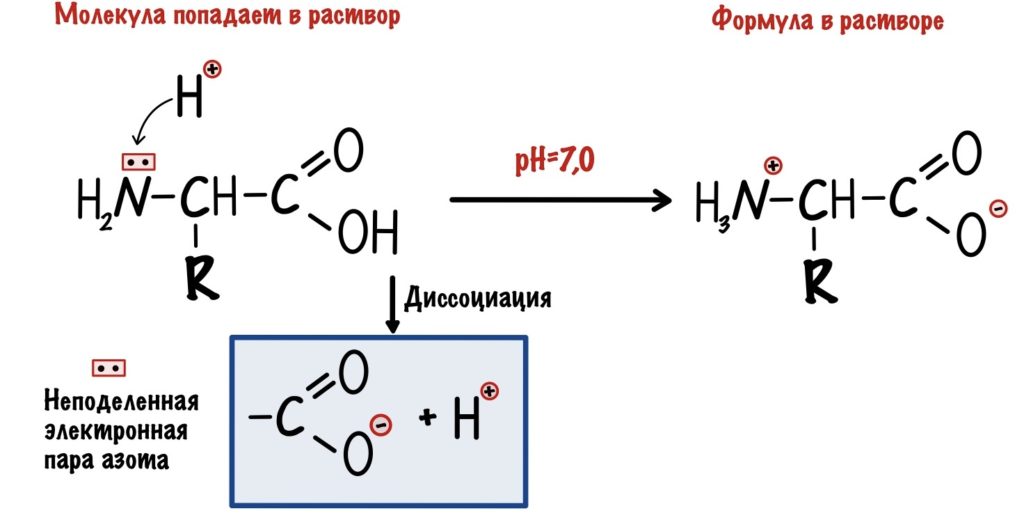
Еще разок, в цитоплазме клетки pH 7,0, то есть в в ней равное количество H+ и OH—. Так как у азота есть неподеленная электронная пара, то он может присоединить протон водорода по донорно-акцепторному механизму. А остаток кислоты диссоциирует с образованием протона. Понятное дело, что радикалы подчиняются таким же правилам.
Полярные радикалы можно разделить на две группы: полярные незаряженные и полярные заряженные.
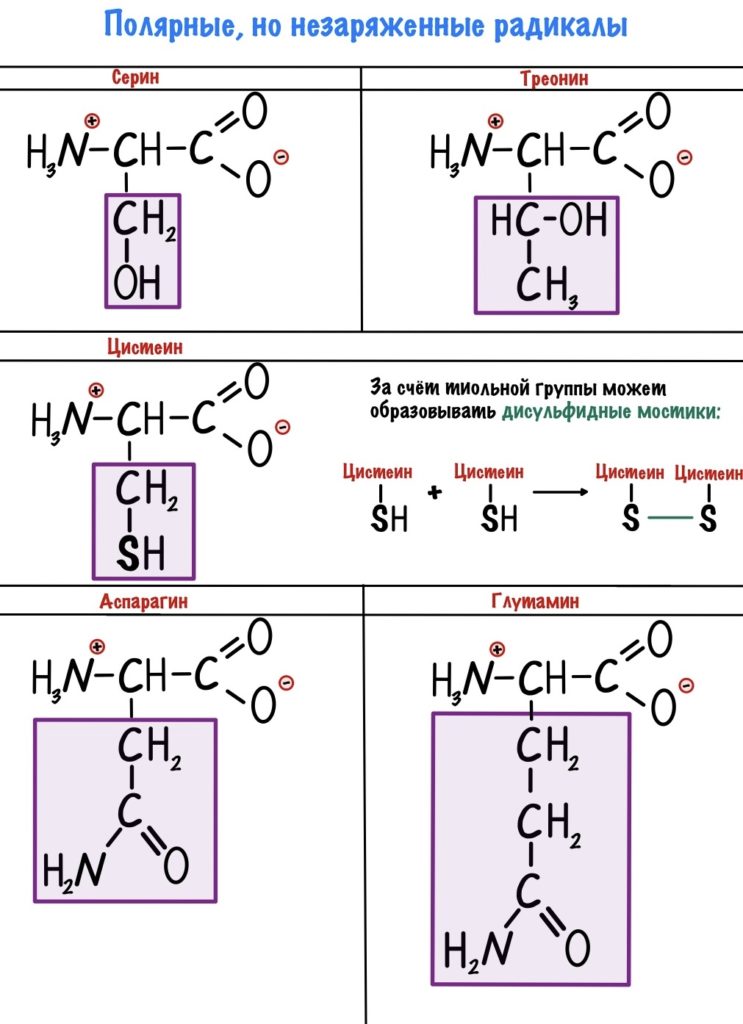
В этих аминокислотах есть сильно электроотрицательные атомы — азот, кислород и сера. С их помощью молекулы образуют водородные связи и растворяются в воде. Но заряда у них нет.
Заряд у радикала может быть положительным или отрицательным, поэтому здесь небольшое деление.
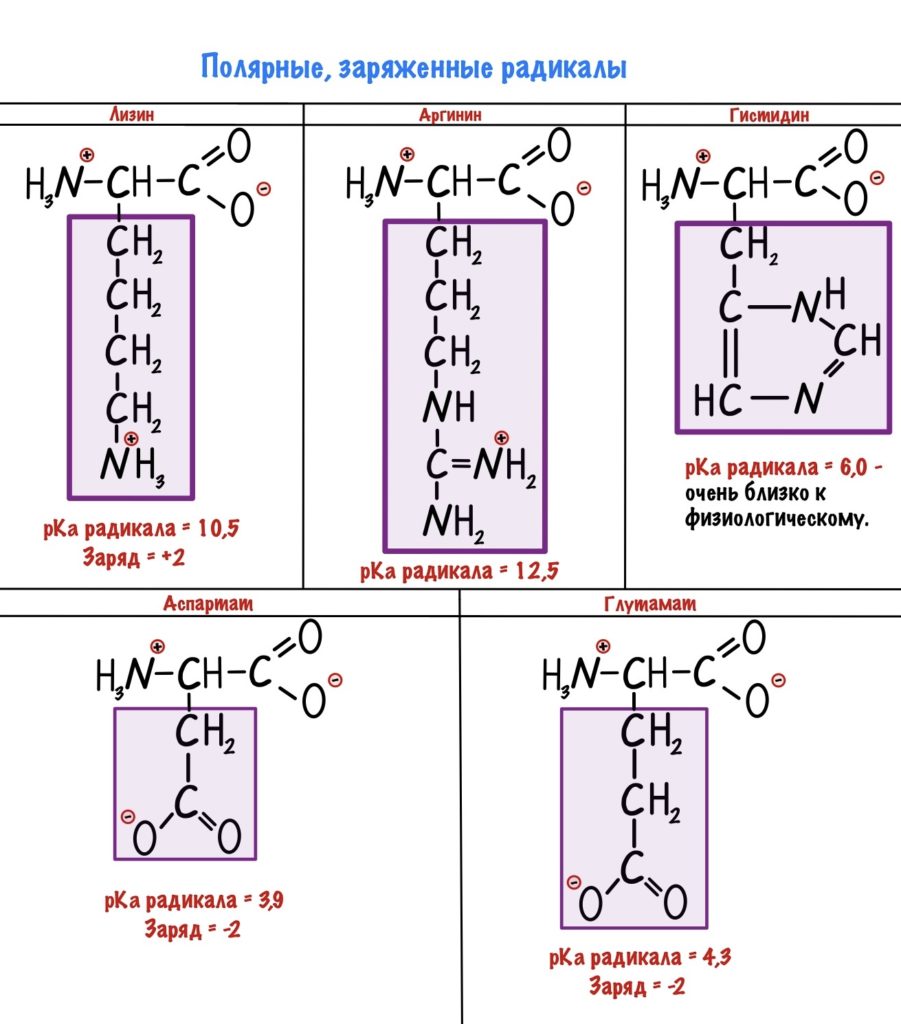
Кстати, лучше растворимы в воде заряженные радикалы. Но разница между полярными заряженными и незаряженными не слишком большая. И еще одно — аспартат и глутамат это название аспарагиновой и глутаминовой кислот в растворе.
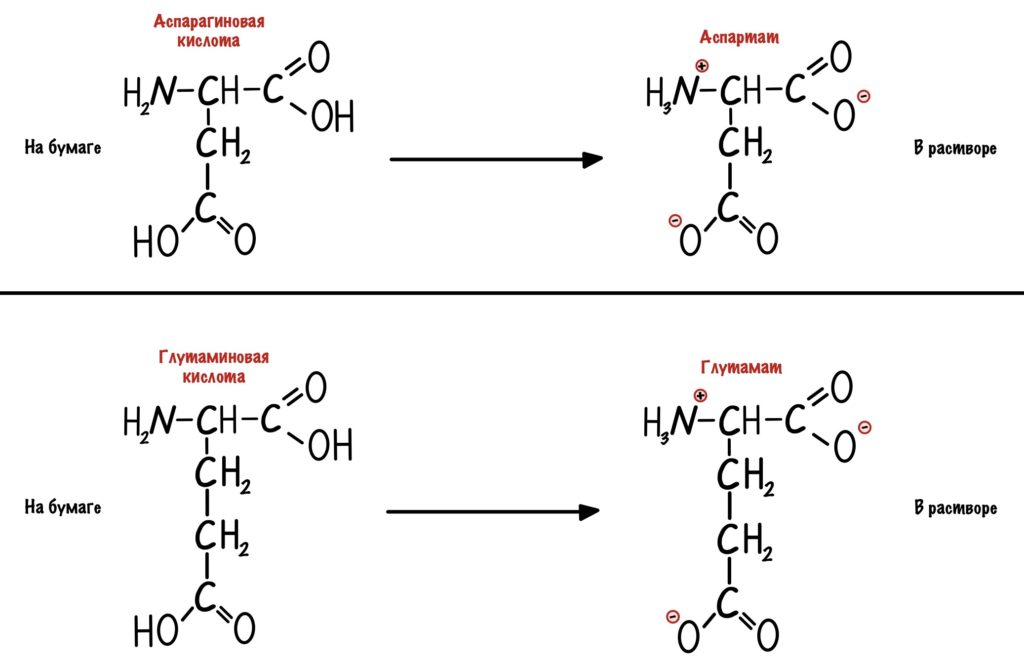
Аминокислоты делятся на полярные и неполярные. Полярные аминокислоты могут быть заряженными или незаряженными.
Аминокислоты называли по месту их выделения или физическим свойствам, поэтому у них такие странные названия. Гликос с греческого — сладкий, вот и глицин сладковат. Так что придется зазубрить это.
Изоэлектрическая точка
Вы уже заметили, что у аминокислот есть положительная и отрицательная части. Не так много молекул имеют такую особенность. Так что аминокислоты — это такой гибрид, поэтому их так и назвали — гибридные ионы. Правда на немецком…. А звучит это так: «Цвиттер-ион». Но как всегда есть один нюанс — у гибридного иона общий заряд молекулы равен нулю.
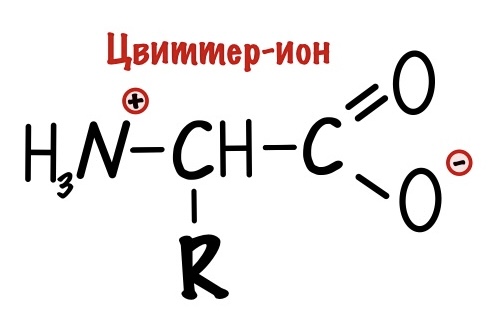
И вы уже смекнули, что не у всех аминокислот будет общий заряд равен нулю. Для неполярных и полярных незаряженных аминокислот это верно, но че делать с заряженными? До этого мы разбирали заряд аминокислот в клетке, то есть при нейтральном pH. Но что будет с ними, если поместить их в другие значения среды, например, в сильнощелочную или кислотную? Аминокислоты будут менять свой заряд и сейчас посмотрим как.
Для начала возьмем что-нибудь простенькое — пусть это будет глицин. Нужно понять, что мы берём раствор — у него будет определённая кислотность среды (pH) равная 7,0. В этом растворе будет множество молекул глицина. Добавляем в этот раствор кислоту. Прямо по капле. Сначала ничего не меняется, но при значении pH=2,34 половина карбоксильных групп глицина присоединит к себе протон водорода. Продолжим уменьшать pH (добавлять кислоту) — всё большее количество глицина присоединяет протон. Что же получается? При снижении pH менее 2,34 большая часть карбоксильных групп глицина присоединит к себе протон. Заряд молекул глицина изменится — теперь он равен +1. Начнем добавлять к этому же раствору щелочь по капле. Дойдем до pH равного 9,6 и от половины аминогрупп глицинов отлетит водород. Заряд молекулы будет -1. Чуть дальше будет хорошая картинка, на которой можно будет проследить за всеми изменениями.
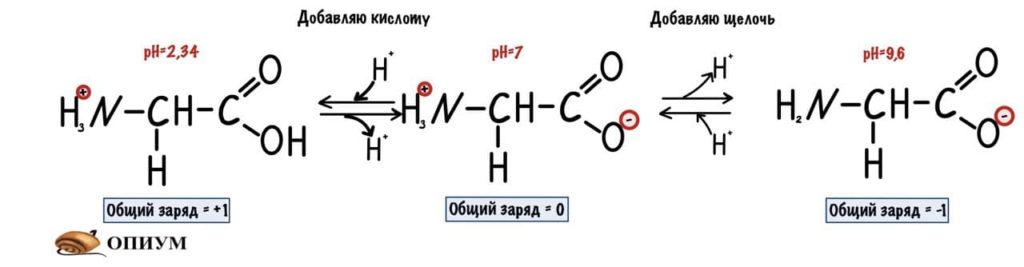
Как же назвать pH при котором происходит переход из одной формы в другую? Очень просто, показатель константы диссоциации или pKa. Константа диссоциации показывает при каком pH среды половина функциональных групп связана с протоном, а другая половина не связана. Получается, что в молекуле глицина pKa карбоксильной группы = 2,34, а pKa аминогруппы = 9,6. Я написал про молекулу глицина, потому что в остальных аминокислотах значения немного отличаются.
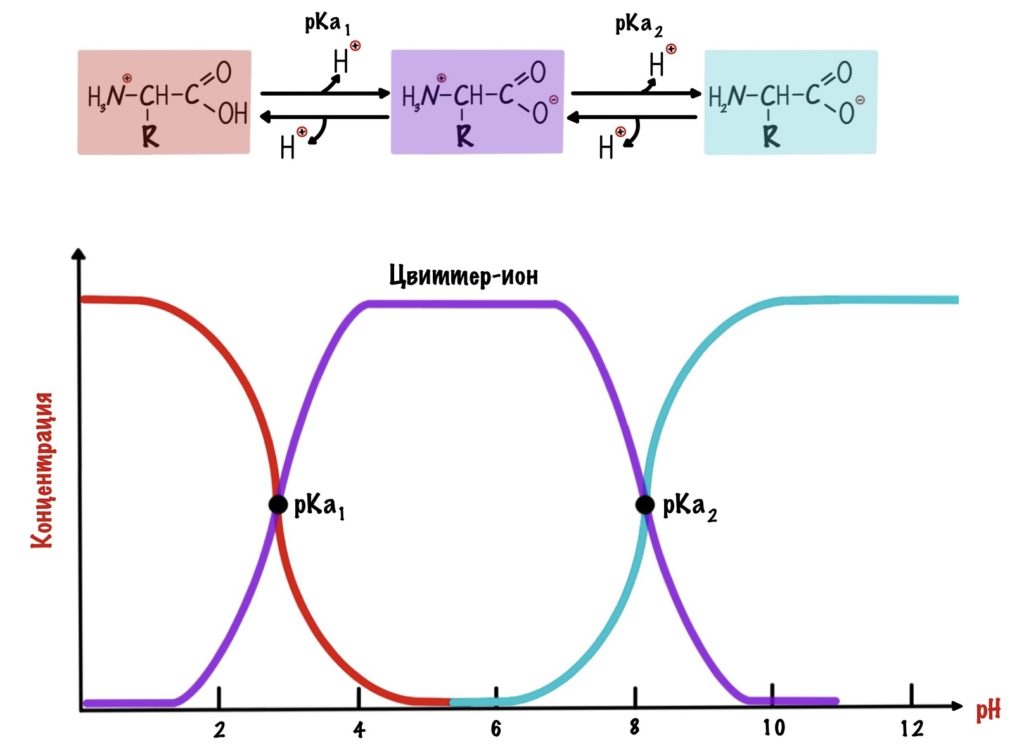
А теперь о том, ради чего все это затевалось — изоэлектрическая точка.
Изоэлектрическая точка — это pH среды, при которой заряд молекулы равен нулю. Да, вот так вот просто. Ее, кстати, можно посчитать — для этого нужно сложить pKa двух ближних функциональных групп и поделить на их количество. А их количество — две.
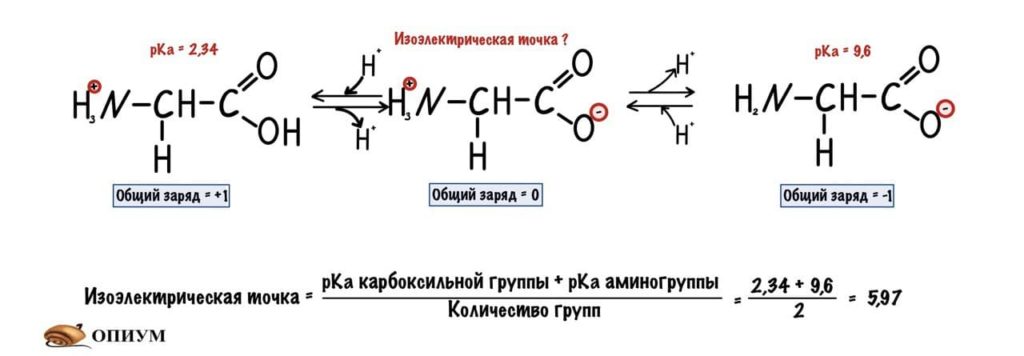
Сделаем тоже самое с молекулами посложнее, начнем с гистидина.
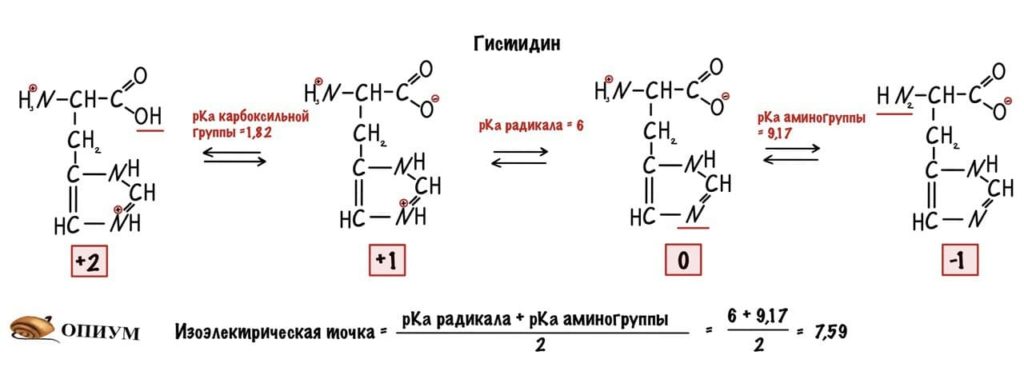
У гистидина есть заряженная группа, поэтому у него побольше вариантов заряда, чем у глицина. Мы видим, что у гистидина карбоксильная группа присоединяет водород при pH =1,82, а аминогруппа отдает протон водорода при pH=9,17. Вот про эти отличия я и говорил до этого, но так-то они не слишком большие. Радикал же отдает протон водорода при pH=6.
Сделаем тоже самое с глутаматом.
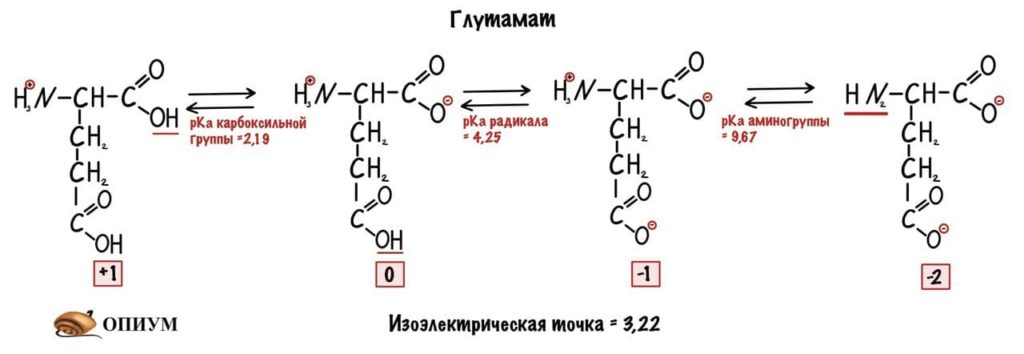
Думаю, что смысл понятен. У каждой аминокислоты своя собственная изоэлектрическая точка. Точки уже давно подсчитаны — достаточно найти их в интернете.
Сделаем красивый вывод:
Любая аминокислота цвиттер-ион, но только в изоэлектрической точке
Зачем это нужно? Ну давайте посмотрим. Мы знаем, что каждая аминокислота несет определенный заряд, но этот заряд меняется от pH среды. Если мы поместим аминокислоты в нейтральную среду и закинем туда катод и анод, то положительно заряженные аминокислоты направятся к аноду, а отрицательные к катоду. Остальные аминокислоты можно будет разделить с помощью изменения pH среды, ведь в изоэлектрической точке у аминокислоты не будет заряда. Нет заряда — нет движения к катоду или аноду, аминокислота стоит на месте. Вот мы и разделили аминокислоты в растворе, можно их изучить.
Образование пептидов
Теперь давайте соединим между собой парочку аминокислот, пусть это будет глицин и аланин. Соединяем их с помощью реакции дегидратации — отщепляем молекулу воды и получаем пептид.
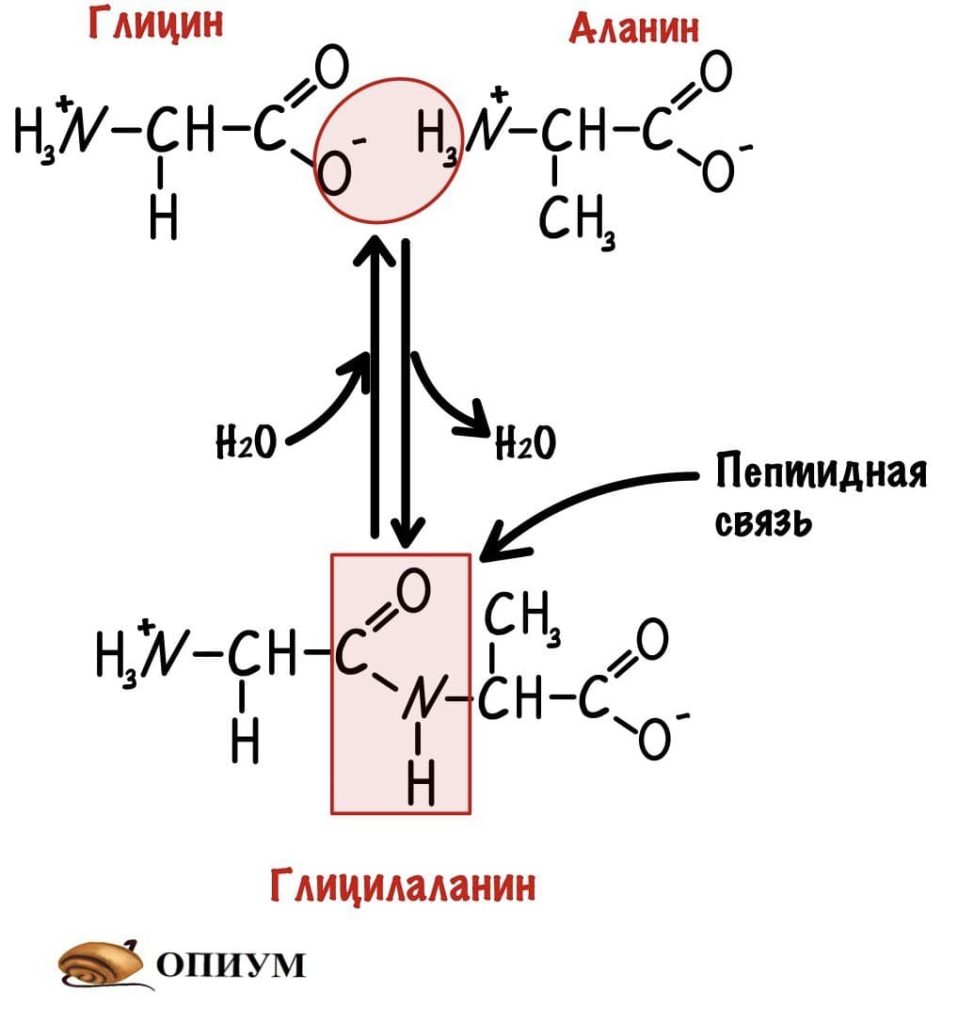
Какие группы вступали в реакцию? Да, аминогруппа и карбоксильная группа. Получается, что пептидная связь — это связь между аминогруппой одной аминокислоты с карбоксильной группой другой аминокислоты. Так как соединены две аминокислоты, то название молекулы — дипептид. Ничего не мешает мне присоединить еще одну.
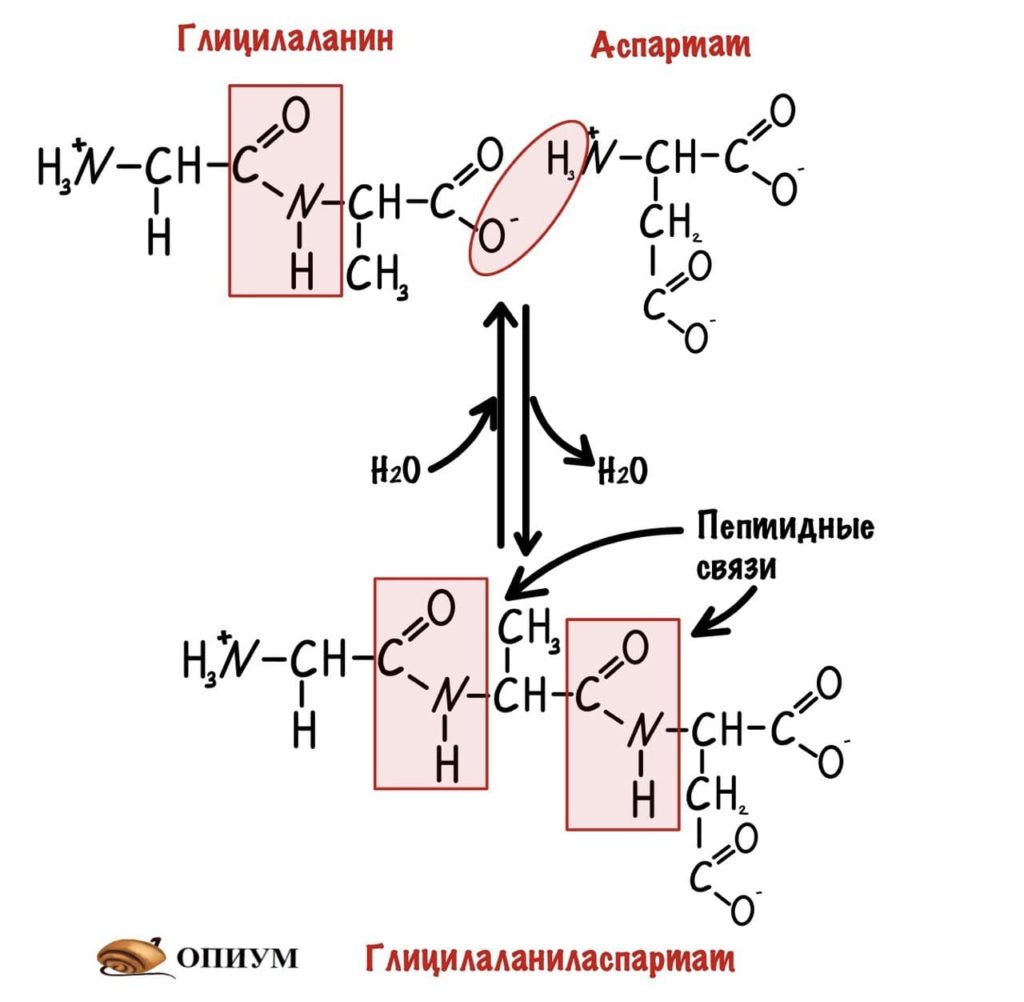
И это уже трипептид. Если соединены до 10 пептидов, то это олигопептид. От 10 до 50 — полипептид, ну а если больше 50, то это белок. Как видите реакция обратима, можно провести гидратацию по пептидной связи и пептид разрушится. На самом деле реакция гидратации идет намного лучше, а вот для дегидратации нужен источник энергии — АТФ, и рибосомальная РНК. Так что для синтеза пептидов/белков организм неплохо так тратится.
Ну и вы заметили, что я располагаю радикалы с разных сторон — то сверху, а то снизу. Это транс положение, оно более устойчиво, но можете писать как хотите.
Белок — это пептид, который содержит более 50 остатков аминокислот
Пептидная связь
У пептидной связи есть свои секретики, но мы не дадим ей хранить их просто так. Главный секрет в том, что двойная связь находится не у кислорода, а у азота… Хотя это не совсем двойная связь, но близка к ней. Как же это происходит? У азота есть неподеленная электронная пара, электроны могут перейти от азота к кислороду, а двойная связь перейдет от кислорода к азоту — неплохой такой обменчик. Это явление называется резонанс пептидной связи, именно из-за него во всех учебниках пишут про «частично-двойной характер пептидной связи».
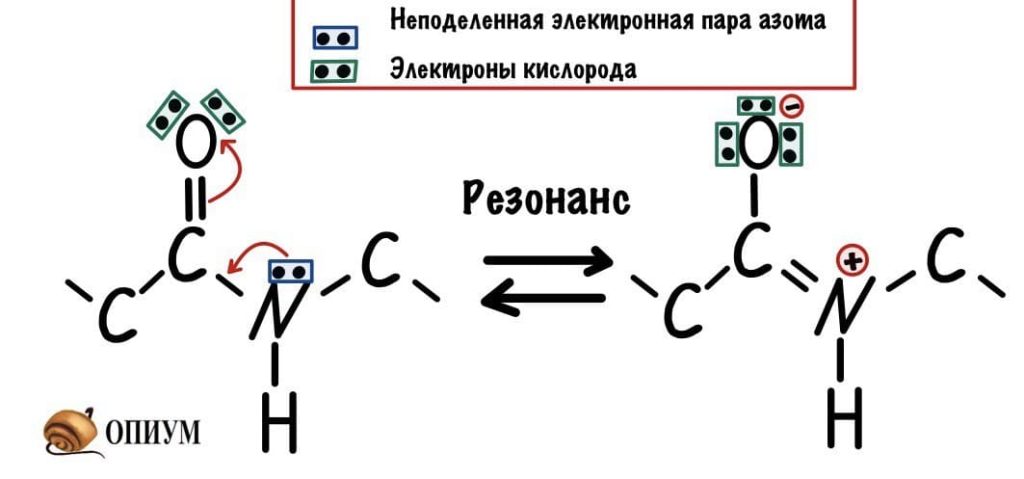
Пойдем еще немного дальше, о чем нам говорит двойная связь? Правильно, о гибридизации углерода — она здесь sp2. А значит угол между связями углерода 120 градусов, здесь они не прямо 120 градусов, но близки к этому. Идем дальше. Азот здесь тоже в sp2 гибридизации, понятно какие углы и у него. Но к чему я это все? Ах да, как будет выглядеть молекула?
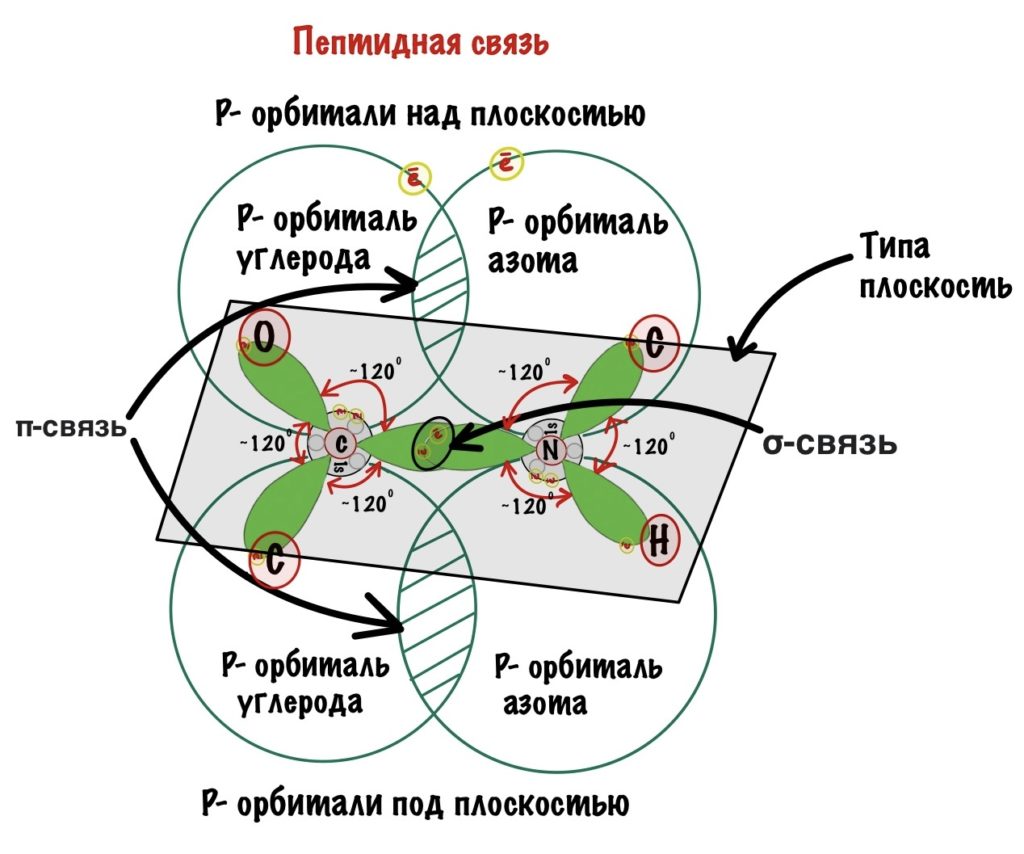
Так как все углы по 120 градусов, то все 6 атомов — 3 углерода, азот, водород и кислород, лежат в одной плоскости, как будто на ладошке. За счет того, что углерод и азот образуют две связи — одну пи и одну сигму, вращение вокруг этих связей практически невозможно. Но об этом чуть позже, сейчас давайте упростим эту схему.
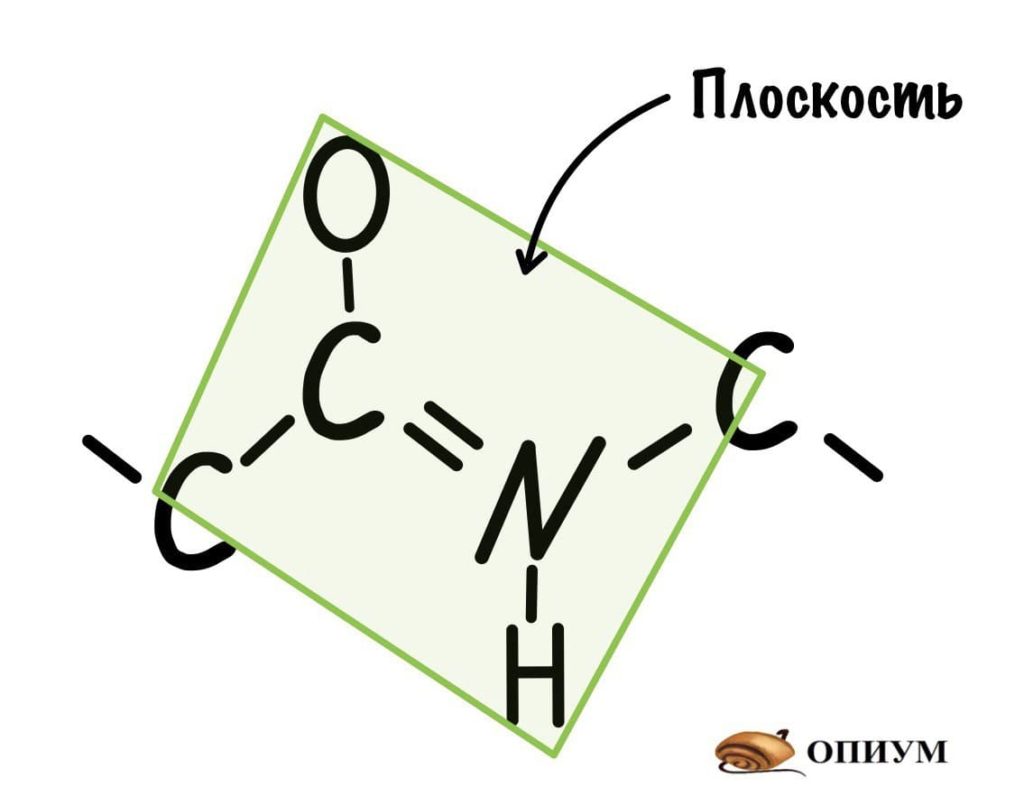
Это мы сделали только с одной пептидной связью, но что если добавить вторую? Получится кое что интересненькое…
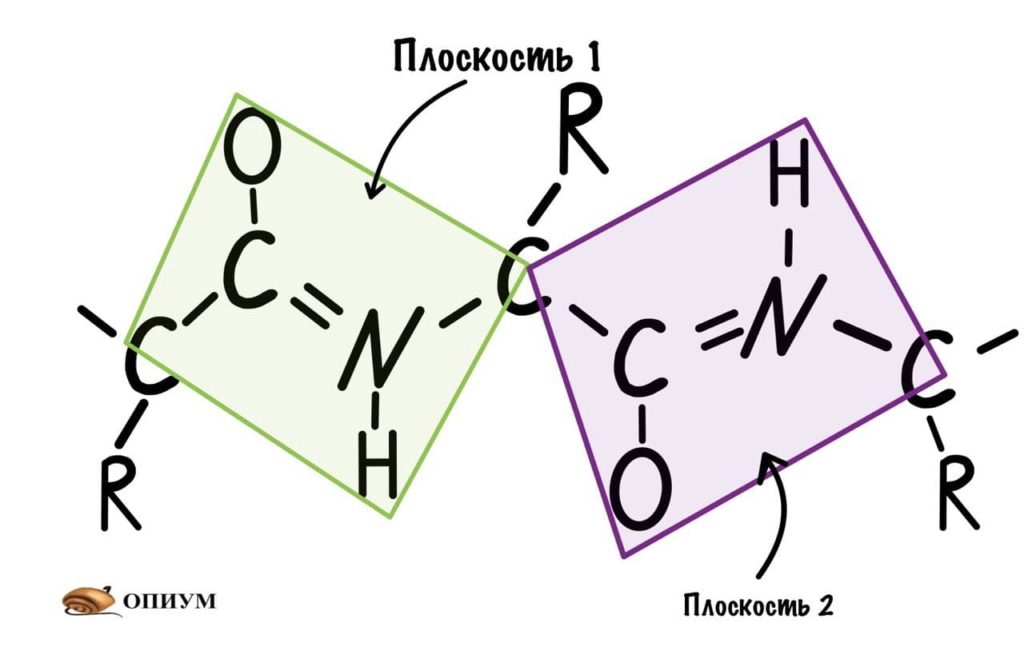
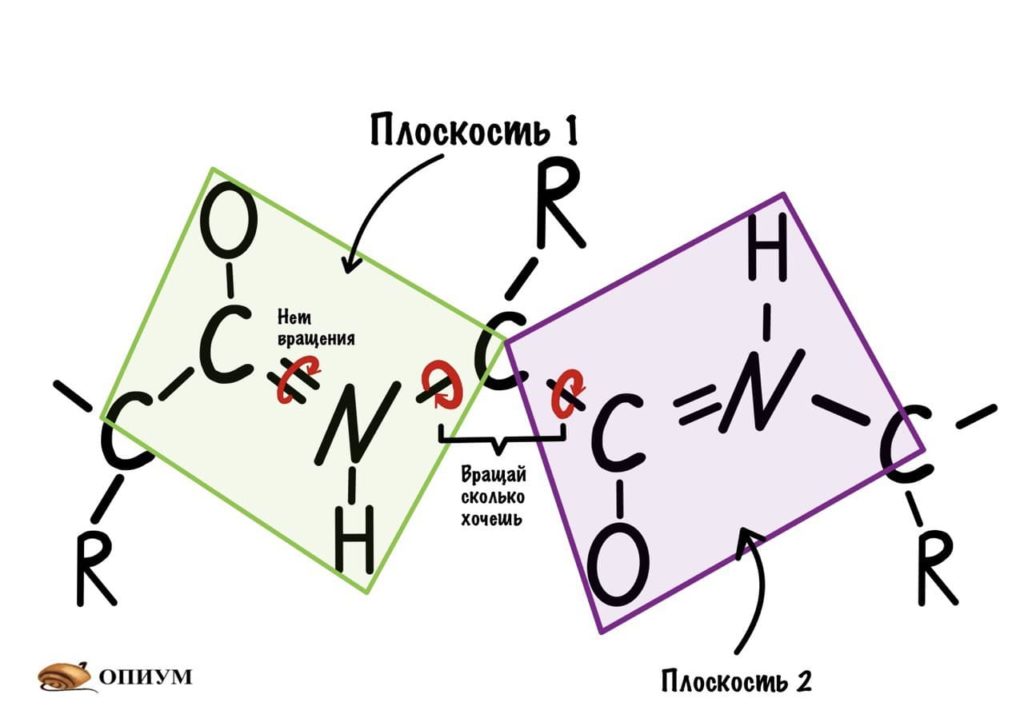
Следующая пептидная связь такая же, как и предыдущая. Получается, что опять 6 атомов лежат в одной плоскости, вы видите, что один атом углерода принадлежит сразу к двум плоскостям и это удивительно! Можно даже подумать, что все эти пептидные связи будут лежать в одной и той же плоскости, но это не так, а виной этому — вращение вокруг связей.
Диэдральные или торсионные углы
Название пугающее, но сейчас как устроим этим углам! Так, мы уже говорили о том, что вокруг пептидной связи не повращаться из-за того, что она частично двойная. Но ведь есть и другие связи, вокруг которых можно устроить веселуху.
Понимаю, что представить это не так уж и легко, но можно попробовать сделать! Получится конечно не совсем так, но принцип поймем. Возьмем ручку и два колпачка, засунем бумажку под каждый колпачок и начнем крутить. Условимся, что мои пальцы — альфа-углеродный атом, то есть место пересечения двух плоскостей.



Теперь мы поняли, как происходит вращение, но это еще не все. Существуют определенные углы между плоскостями и всего их два. Представьте, что нам захочется найти угол между углеродами, у которых карбоксильная группа, двух плоскостей. Или угол между двумя атомами азота, опять же, двух разных плоскостей. Задачка кажется сложной… Но перед этим, а зачем я вообще мучаю вас этим? Дело в том, что когда мы дойдем до конформации белковых молекул, то благодаря этим углам мы поймем: как и почему образуется альфа-спираль, тоже самое с бета-складчатостью. Так что потерпите немного!
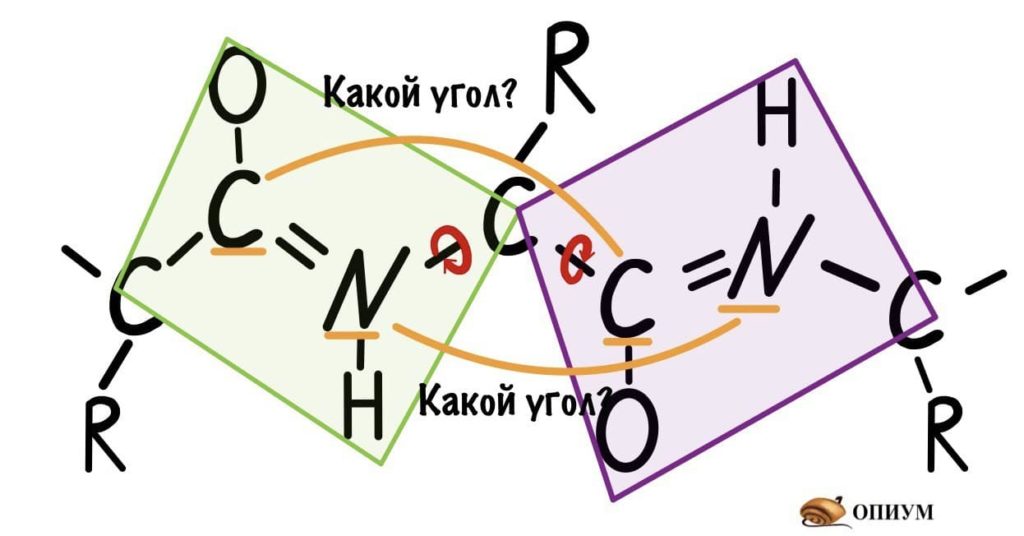
Если посмотреть на эту схему, то можно кое-что прикинуть: если мы будем вращать связь между N и C, то углерод с карбоксильной группой изменит положение относительно углерода другой плоскости, а вот азот останется на том же месте — угол между двумя азотами не изменится. А вот если начнем вращать связь между C и C, то все будет наоборот: угол между азотами изменится, но вот углероды с карбоксильной группой останутся на месте. Сложновато, но чуть дальше я дам пространственную картинку. Пока что мы пришли к выводу, что связь между N и C влияет на угол между углеродами — этот угол называется фи. А вот связь между C и C влияет на угол между атомами азота — угол пси.
Теперь можно и добавить атомы водорода в схему, они скоро нам понадобятся.
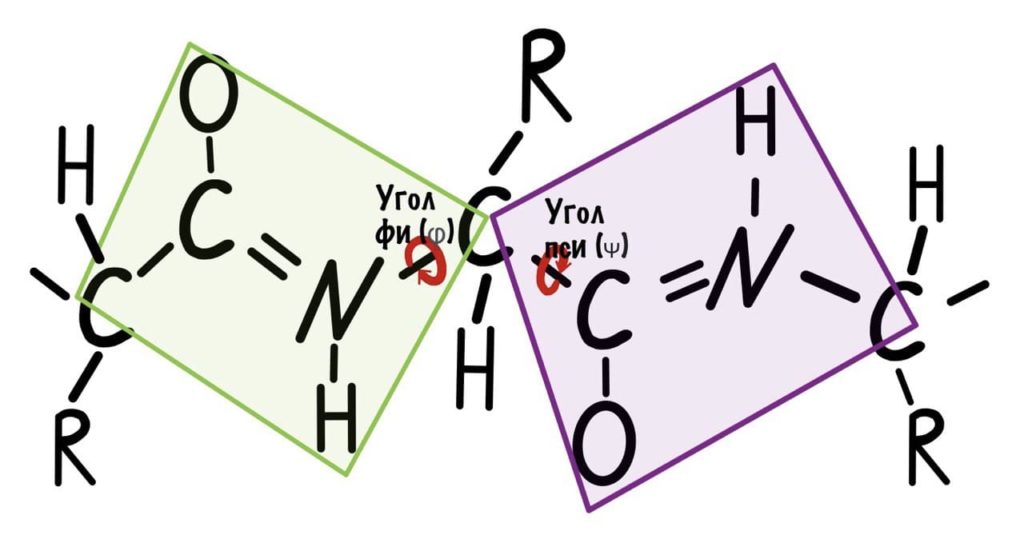
Добавим реалистичности, центральным радикалом у нас будет -CH3, а остальные радикалы уберем.
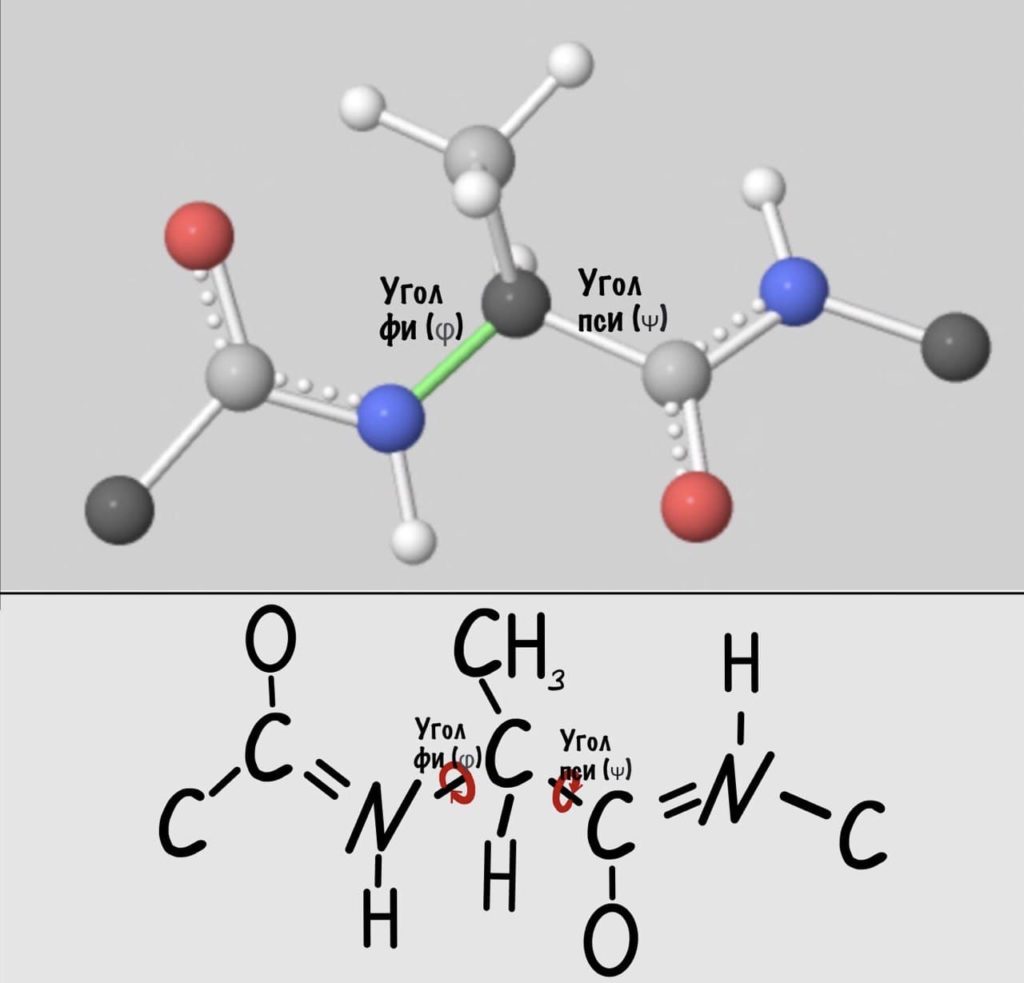
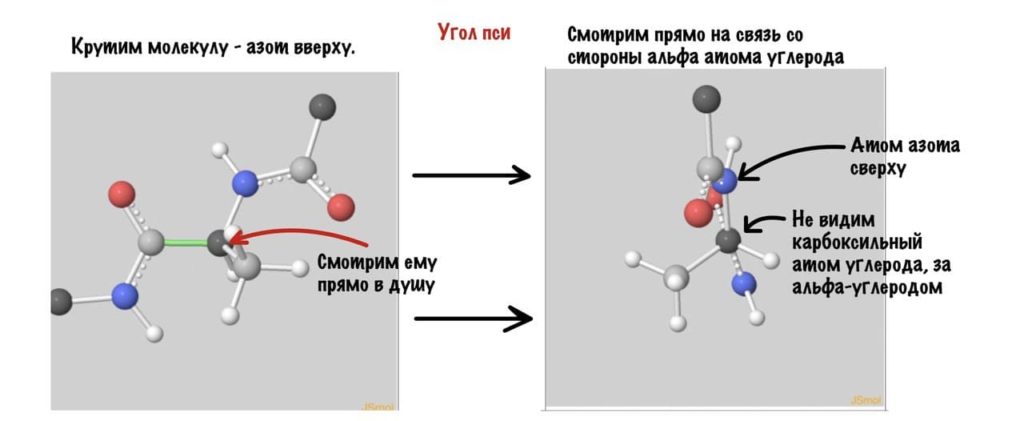
А теперь главный вопрос — как измерить эти углы? Хорошо, что уже это придумали… И мы можем сделать это вместе — заходите сюда и поехали! Первым делом нам нужно перевернуть молекулу так, чтобы расположить атом углерода с карбоксильной группой сверху. Зачем такие выкрутасы? Расскажу позже. А теперь посмотрим прямо в альфа атом углерода, да так что за ним спрятался азот. Как-то это странно звучит, но давайте попробуем.
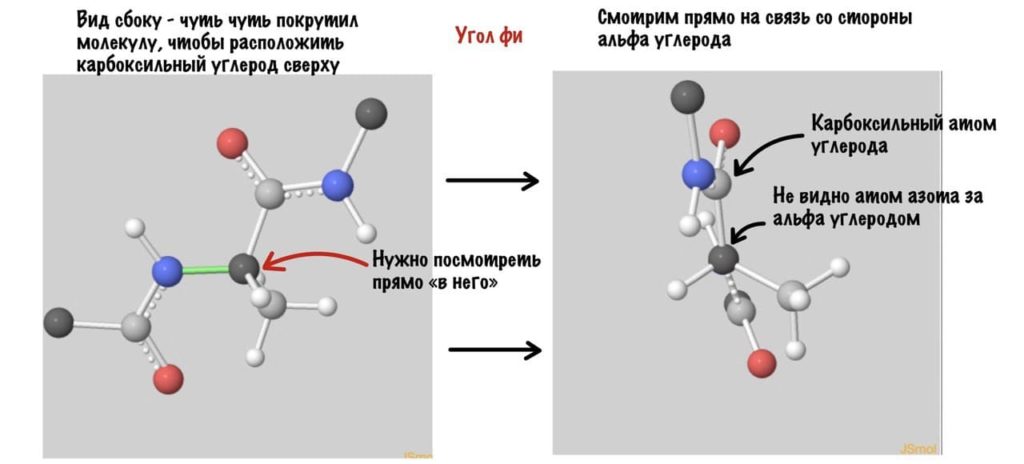
Еще это можно посмотреть графически с помощью проекций Ньюмана.
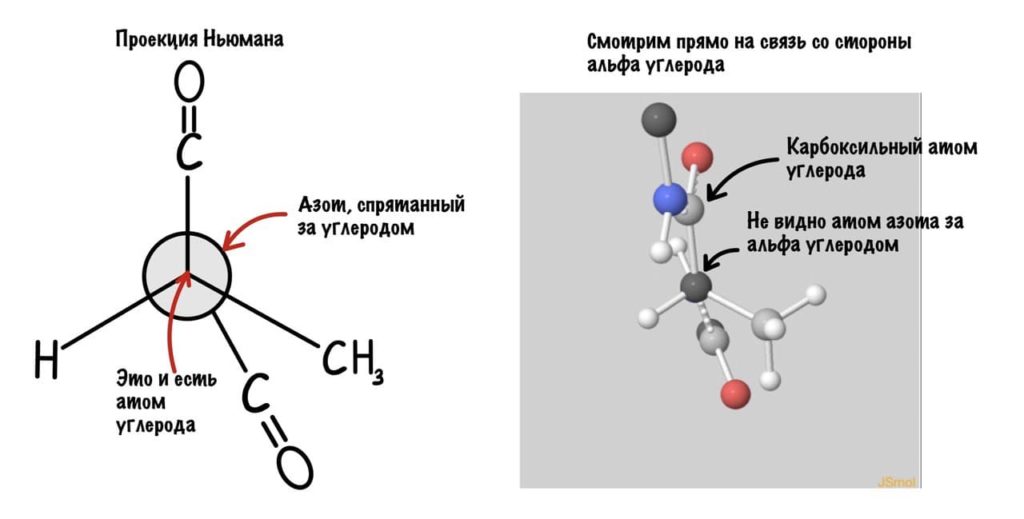
Так, повторим что такое угол фи — это угол между двумя карбоксильными атомами углерода. На рисунке уже их видно.
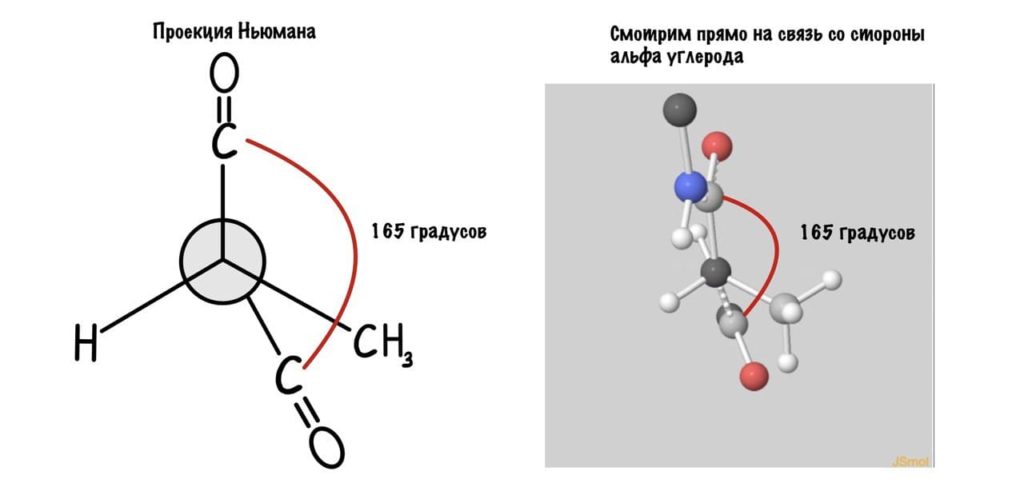
Поняли зачем так крутили молекулу? Да, просто так нам удобнее смотреть угол. А теперь начнем вращать и посмотрим как меняются углы.
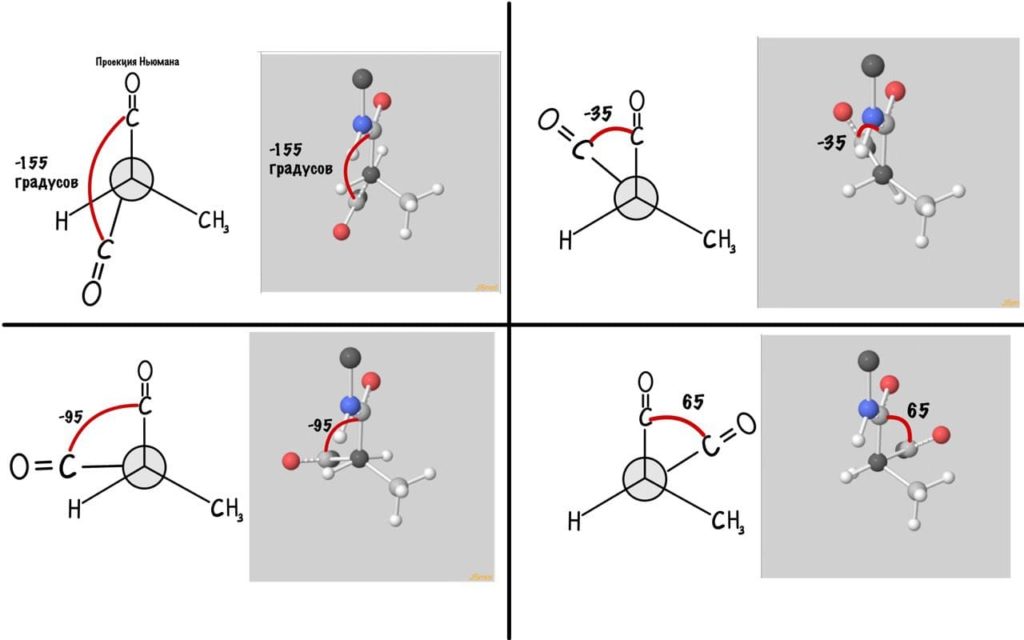
Угол пси по такой же логике. Крутим молекулу, чтобы атом азота оказался сверху и смотрим прямо в альфа атом углерода.
Еще разок построим проекцию Ньюмана, она немного отличается, и сразу же отметим углы.
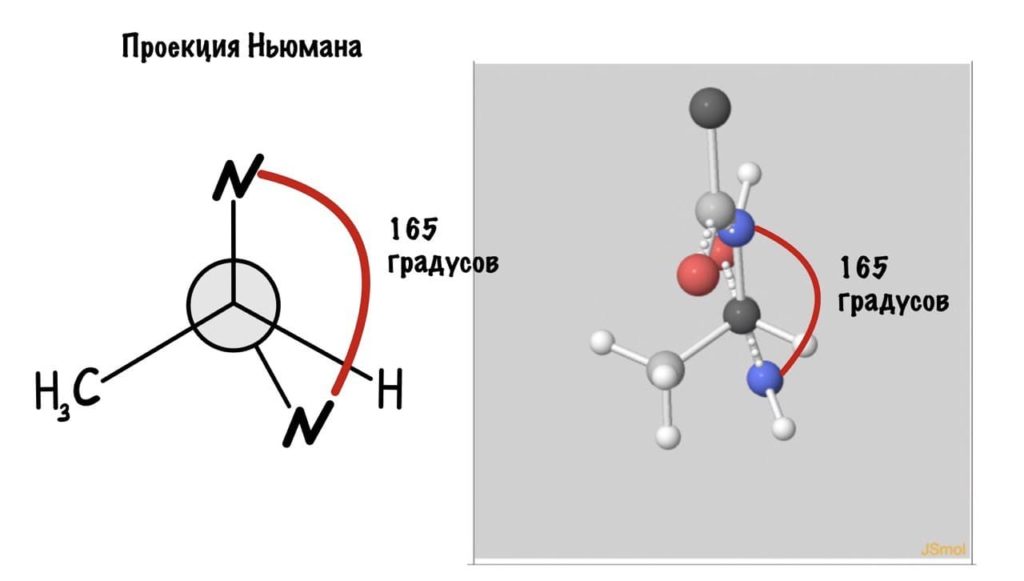
Думаю, что принцип понятен. Дальше можете покрутить сами, правильно? Я не сказал про одно большое «НО» — не каждый угол возможен, так как у атомов есть электронные оболочки, которые заряжены отрицательно. Если электронные оболочки подходят слишком близко, то они отталкиваются и угол меняется. Какие углы возможны? Для этого еще разок зайдите сюда и включите на панельке справа силы Ван-дер-Вальса и show clashes.
Подробнее о влиянии этих углов в следующей статье.
Пептиды представляют собой короткие полимерные фрагменты, состоящие из аминокислот. Каждый пептид имеет определенную аминокислотную последовательность, обозначенную трехбуквенным или однобуквенным кодом; например, аминокислота аланин сокращенно обозначается как «Ala» или «A.». Заряд пептидов в растворе зависит от кислотности раствора. Изоэлектрическая точка (pI) относится к значению кислотности раствора, при котором пептидная молекула имеет суммарный заряд, равный нулю. Растворимость пептида минимальна в изоэлектрической точке. Используйте доступные веб-серверы для расчета значения pI для пептидной аминокислотной последовательности.
Напишите пептидную последовательность, используя однобуквенный код. Например, если пептид имеет аминокислотную последовательность Ala-Ser-Glu-Leu-Pro (аланин-серин-глюкозная кислота-лейцин-пролин), то последовательность из одной буквы “ASELP”. При необходимости обратитесь к таблице перевода из трех букв в одну, приведенную в разделе «Ресурсы».
Используйте любой веб-браузер, Internet Explorer или Firefox, например, чтобы перейти на сервер, который вычисляет изоэлектрическую точку пептида (pI); см. Ресурсы.
Введите пептидную однобуквенную последовательность – «ASELP» в нашем примере – в поле и нажмите «Compute».
Считайте значение изоэлектрической точки (pI), указанное в строке «Теоретическое значение pI / Mw». В нашем примере pI составляет 4, 00. Обратите внимание, что сервер также рассчитывает молекулярную массу (Mw) пептида.
 пептидов. Биохимия.")
Смотреть видео:
#химия #химияпросто #неорганика #егэпохимии #эксперименты #химик #егэхимия #опыты #химияегэ
Свежая информация для ЕГЭ и ОГЭ по Химии (листай):
С этим видео ученики смотрят следующие ролики:

Задача на определение формулы трипептида. Биохимия.
День знаний

Движение пептидов в электрофоретическом поле при разных рН. Биохимия.
День знаний

Степень окисления. Определение степеней окисления. Часть 1
Your system education

Олимпиадная задача на определение состава смеси органических веществ.
День знаний
Облегчи жизнь другим ученикам – поделись! (плюс тебе в карму):
10.12.2021
Молекулы белков представляют собой амфотерные основания, поскольку содержат свободные амино- и карбоксигруппы. При рН раствора меньше 7 они имеют отрицательный заряд, а при рН больше 7 – положительный. При равенстве данных зарядов, которое достижимо при установлении в растворе определенной кислотности или щелочности, устанавливается так называемая изоэлектрическая точка белка.
Понятие об изоэлектрической точке
Белки состоят из аминокислот. Некоторые из этих соединений (аргинин, аспарагиновая кислота, гистидин, глутаминовая кислота, лизин) представлены в виде радикалов, содержащих ионогенные группы, то есть такие группы, которые способны к ионизации. Помимо них к ионизации способны альфа-карбоксильная и аминогруппы, расположенные на углеродном и азотном концах полипептидных цепей. Если рН раствора равен 7 или приближен к данной отметке, то в ионизированном состоянии находятся все ионогенные группы. По мере удаления от данного значения рН в ту или иную сторону, причем преимущественно в кислую, белок начинает переход в изоэлектрическое состояние, при котором молекула данного вещества становится электронейтральной, число ионизированных групп стремится к нулю. Величина рН, при которой белки переходят в изоэлектрическое состояние, называется изоэлектрической точкой белков (ИЭТ).

Физико-химическая природа белков
Из-за того, что в состав белков входят карбокси- и аминные группы, они могут диссоциировать как основания и как кислоты. Свободная карбоксильная группа при диссоциации отдает положительно заряженный ион водорода и анион COO-. В результате ион водорода присоединяется к аминогруппе, что характеризует основные свойства белка, в результате чего образуются частицы белка с отрицательным и положительным зарядами. При помещении белка в кислый раствор его кислотная диссоциация будет подавляться из-за значительного присутствия катионов водорода. И наоборот, при помещении его в основный раствор его основная диссоциация будет подавляться из-за присутствия анионов COO-.
Пропускание через белковый раствор электрического тока приведет к тому, что анионы начнут двигаться к катоду, а катионы – к аноду. В любом белке есть определенная величина рН, при которой движение ионов при пропускании тока не будет происходить. В этом случае говорят о равенстве разнонаправленных ионов и равенстве различных степеней (основной и кислой) в белковой молекуле, что характеризуется изоэлектрическим состоянием.
Как известно, вода представляет собой диполь, поэтому она располагает свои частицы вокруг белковой молекулы в зависимости от того, как она заряжена. В изоэлектрической точке молекула белка не имеет гидратно-ориентированной оболочки. Если осуществляется осаждение белков, необходимо, прежде всего, разрушить гидратную оболочку, сняв электрический заряд.
Использование ИЭТ в промышленности
В изоэлектрическом состоянии некоторые свойства раствора белка, такие как набухание, вязкость, осмотическое давление, светопропускание, имеют минимальные значения, при этом показатель преломления и оптическая плотность достигают, наоборот, максимальной величины. Изоэлектрическую точку белка можно определить опытным путем, определяя зависимость указанных выше свойств белкового раствора: от величины рН, при этом по положению экстремумов на графиках определяют ИЭТ. В изоэлектрическом состоянии казеин способен осаждаться, что применяется при производстве сыров и кисломолочных продуктов, для получения казеина из обезжиренного молока, как сырья в различных производствах (казеиновые клеи, искусственные продукты питания и т. д.). Измерение ИЭТ позволяет оценить качество белка, в частности, молочного продукта на наличие примесей. Это актуально на сегодняшний день, поскольку введение растительных добавок в молочную основу позволяет заменить часть животного белка растительным, который является более дешевым.
Помимо этого, изоэлектрическая точка белка может использовать при очистке сточных вод от птицефабрик. Так, основная доля загрязнений сточных вод убойного цеха птицефабрики приходится на белки крови. Учитывая то, что ИЭТ большинства белков находится в зоне слабокислой реакции среды, наиболее полное извлечение белков будет происходить при слабокислой реакции среды при величине рН, стремящейся к ИЭТ.
Факторы, оказывающие влияние на ИЭТ

На ИЭТ оказывает влияние несколько факторов. Рассмотрим, от чего зависит изоэлектрическая точка белков. Прежде всего, она определяется преобладанием аминных или карбоксигрупп в составе молекулы белка. Большая часть белков представляют собой более сильные кислоты по сравнению с основаниями, поэтому для них ИЭТ меньше 7. Имеется группа белков, которые являются более сильными основаниями, чем кислотами, для них ИЭТ больше 7. Установлена сильная корреляционная зависимость между изоэлектрической точкой белка и содержанием ионов солей в растворе. Концентрация белка не оказывает никакого влияния на данный показатель. Рассмотренные факторы позволяют понять, почему изоэлектрическая точка различна для разных белков.

Примеры ИЭТ белков:
- пепсин имеет значение ИЭТ около 1;
- казеин и желатин – 4,7;
- яичный альбумин – 4,8;
- муцин – 2,7;
- пепсиноген – 3,7;
- альбумины – 4,6;
- инсулин – 5,3;
- оксигемоглобин – 6,8;
- карбоксигемоглобин – около 6,9;
- миоглобин – 7,0;
- химотрипсин – 8,6;
- цитохром С – 10,5;
- сальмин – 12.
ИЭТ и ее определение

Все методы определения изоэлектрической точки белков основаны на приготовлении буферных растворов, имеющих отличающуюся реакцию среды. Во все эти растворы помещаются одинаковые навески изучаемого белка, который может быть как в сухом виде, так и в виде раствора. Используются различные методы определения ИЭТ. Как определить изоэлектрическую точку белка?
Основными методами определения ИЭТ являются электрофорез, по минимуму вязкости и связанный с применением водоотнимающих веществ. Могут использоваться и некоторые другие методы, такие, как определение по степени набухания сухого белка, скорости застудевания, но они менее точные и требуют наличия большого количества белка.
Электрофорез
При использовании данного метода в прибор для его осуществления помещаются полоски хромотографической или фильтровальной бумаги, смоченные определенным буферным раствором. Посередине каждой полоски делается карандашная отметка, в которую при помощи пипетки наносится одна капля изучаемого раствора белка. Затем прибор включают и через эти полоски пропускают электрический ток. Макромолекулы изменяют свой заряд в зависимости от величины рН буферного раствора. Если величина рН превышает ИЭТ, то наблюдается отрицательный заряд макромолекул, и наоборот.

Если рН равно ИЭТ, то макромолекулы становятся нейтрально заряженными. Через определенное время подача тока прекращается, полоски бумаги достаются из прибора и высушиваются, после чего пятна белка опрыскивают нингидрином для их проявления. ИЭТ устанавливают по буферному раствору полоски бумаги, где белковое пятно осталось там же, где была нанесена капля. При необходимости этот метод может быть применен и для тонкого фракционирования белков.
Применение других методов для определения ИЭТ
При нахождении в изоэлектрическом состоянии молекулы белков менее гидратированы, поэтому изоэлектрическую точку белка можно определить, используя метод по минимуму вязкости. Для его применения необходимо наличие вискозиметра. С помощью этого прибора определяют относительную вязкость буферных растворов. Молекулы белка, находящегося в изоэлектрическом состоянии, свернуты, поэтому самая небольшая вязкость будет у раствора, в котором его рН будет совпадать с ИЭТ.
На этом же свойстве основан метод, связанный с действием водоотнимающих средств. В качестве таких средств могут выступать ацетон, эфир или спирт. Выделение белков из соответствующих растворов потенциально происходит тем быстрее и полнее, чем полнее соответствует реакция среды ИЭТ. В изоэлектрической точке растворы белков неустойчивы.
Таким образом, существуют различные методы определения изоэлектрической точки белка. И ее определение должно выполняться в зависимости от имеющегося оборудования, материалов, количества белка.
Устойчивость белка в ИЭТ
В изоэлектрической точке белка силы отталкивания между белковыми частицами в макромолекуле ослабевают, благодаря чему происходи агрегация этих молекул и белок выпадает в осадок. Это свидетельствует о том, что в ИЭТ белок неустойчивый за счет потери заряда, который является фактором стабилизации водных белковых растворов. Если к белку добавить кислоту или основание, то молекулы перезаряжаются, белок осуществляет переход в раствор.

В заключение
Таким образом, изоэлектрическая точка белка представляет собой значение реакции среды (рН), при котором в белковой молекуле отмечается равенство разнонаправленных (отрицательных и положительных) зарядов и равенство различных степеней (основных и кислотных) диссоциации. В данной точке белок теряет заряды и становится неустойчивым, вследствие чего выпадает в осадок. Молекула белка сворачивается, в то время, когда она несет в себе определенные заряды, она распрямлена в виде нити.
Лекция
№ 47
АМИНОКИСЛОТЫ. ПЕПТИДЫ
План
- Методы получения.
- Химические свойства.
- Аминокислоты, входящие в состав
белков. - Пептиды
Аминокислоты
– гетерофункциональные соединения, содержащие карбоксильную и аминогруппы. По
взаимному расположению функциональных групп различают a -,b -, g – и т.д. аминокислоты.
Аминокислоты, содержащие аминогруппу на конце цепи, называют w -аминокислотами.

1. Методы получения
!) Аммонолиз галогензамещенных кислот.

Метод используется для синтеза a
-аминокислот из доступных a -галогензамещенных
кислот.
2) Метод Штеккера- Зелинского
Включает стадии образования аминонитрила при взаимодействии альдегида с HCN
и NH3 c последующим гидролизом его в аминокислоту. В качестве
реагента применяют смесь NaCN и NH4Cl.

Метод применим для синтеза только a
-аминокислот.
3) Алкилирование N-фталимидмалонового эфира

4) Присоединение аммиака к a
,b -непредельным карбоновым
кислотам.

Метод применим для синтеза b
-аминокислот.
5) Из оксимов циклических кетонов перегруппировкой Бекмана.

Метод используется для синтеза w
-аминокислот.
2. Химические свойства
Аминокислоты
дают реакции, характерные для карбоксильной и аминогрупп, и, кроме того,
проявляют специфические свойства, которые определяются наличием двух
функциональных групп и их взаимным расположением.
2.1. Кислотно-основные свойства
Аминокислоты содержат кислотный и основный центры и являются амфотерными
соединениями. В кристаллическом состоянии они существуют в виде внутренних
солей (биполярных ионов), которые образуются в результате внутримолекулярного
переноса протона от более слабого основного центра (СОО–) к более
сильному основному центру (NH2).

Ионное строение аминокислот подтверждается их физическими свойствами.
Аминокислоты – нелетучие кристаллические вещества с высокими температурами
плавления. Они нерастворимы в неполярных органических растворителях и
растворимы в воде. Их молекулы обладают большими дипольными моментами.
Форма существования аминокислот в водных растворах зависит от рН. В кислых
растворах аминокислоты присоединяют протон и существуют преимущественно в виде
катионов. В щелочной среде биполярный ион отдает протон и превращается в анион.

При некотором значении рН, строго определенном для каждой аминокислоты, она
существует преимущественно в виде биполярного иона. Это значение рН называют изоэлектрической
точкой (рI). В изоэлектрической точке аминокислота не имеет
заряда и обладает наименьшей растворимостью в воде. Катионная форма
аминокислоты содержит два кислотных центра (COOH и NH3+)
и характеризуется двумя константами диссоциации рКа1 и рКа2.
Значение рI определяется по уравнению:
![]()
2.2. Реакции по аминогруппе
Дезаминирование
Аминокислоты содержат первичную аминогруппу и подобно первичным аминам
взаимодействуют с азотистой кислотой с выделением азота. При этом происходит
замещение аминогруппы на гидроксильную.
RCH(NH2)COOH + HNO2 ®
RCH(OH)COOH + N2 + H2O
Реакция используется для количественного определения аминокислот по объему
выделившегося азота (метод Ван-Слайка).
Алкилирование и арилирование
При взаимодействии аминокислот с избытком алкилгалогенида происходит
исчерпывающее алкилирование аминогруппы и образуются внутренние соли.

Аминокислоты арилируются 2,4-динитрофторбензолом (ДНФБ) в щелочной среде.
Реакция протекает как нуклеофильное замещение в активированном ароматическом
кольце.

Реакция используется для установления аминокислотной последовательности в
пептидах.
Ацилирование
Аминокислоты взаимодействуют с ангидридами и хлорангидридами с образованием
N-ацильных производных.

Реакция используется для защиты аминогруппы в синтезе пептидов. Такая защита
должна легко сниматься, а амиды, как известно, гидролизуются в жестких
условиях. При разработке методов синтеза пептидов были найдены защитные группы,
которые легко удаляются путем гидролиза или гидрогенолиза.
Карбобензоксизащита:

трет-Бутоксикарбонильная защита (БОК-защита).

Легкость снятия защиты обусловлена устойчивостью бензил- и трет-бутил-катионов,
которые образуются в качестве интермедиатов.
2.3. Реакции по карбоксильной группе
Декарбоксилирование
При сухой перегонке в присутствии гидроксида бария аминокислоты
декарбоксилируются с образованием аминов.

Этерификация
Аминокислоты взаимодействуют со спиртами в присутствии газообразного HCl как
катализатора с образованием сложных эфиров.

В отличие от самих аминокислот, их сложные эфиры – легко летучие соединения
и могут быть разделены путем перегонки или газожидкостной хроматографии, что
используется для анализа и разделения смесей аминокислот, полученных при
гидролизе белков.
Получение галогенангидридов и ангидридов
При действии на защищенные по аминогруппе аминокислоты галогенидов фосфора
или серы образуются хлорангидриды.

Реакция используется для активации карбоксильной группы при нуклеофильном
замещении. Чаще для этой цели получают смешанные ангидриды, которые являются
более селективными ацилирующими реагентами.

Реакция используется для активации аминогруппы в синтезе пептидов.
2.4. Специфические реакции аминокислот
Реакции с одновременным участием карбоксильной и аминогрупп идут, как
правило, с образованием продуктов, содержащих термодинамически устойчивые 5-ти-
и 6-тичленные гетероциклы.
Комплексообразование
a -Аминокислоты образуют прочные
хелатные комплексы с ионами переходных металлов (Cu, Ni, Co, Cr и др.).

Отношение аминокислот к нагреванию
Превращения аминокислот при нагревании зависят от взаимного расположения
карбоксильной и аминогруппы и определяются возможностью образования термодинамически
стабильных 5-ти- 6-тичленных циклов
a -Аминокислоты вступают в реакцию
межмолекулярного самоацилирования. При этом образуются циклические амиды – дикетопиперазины.

b -Аминокислоты при нагревании
переходят a ,b -непредельные кислоты.

g – и d
-Аминокислот претерпевают внутримолекулярное ацилирование с образованием
циклических амидов – лактамов.

Нингидриновая реакция
При взаимодействии a -аминокислот с
трикетоном – нингидрином происходит одновременное окислительное дезаминирование
и декарбоксилирование с образованием альдегида и окрашенного продукта
конденсации.

Реакция используется для количественного анализа аминокислот методом
фотометрии.
- a -Аминокислоты,
входящие в состав белков
3.1.
Строение и классификация
Природные аминокислоты отвечают общей формуле RCH(NH2)COOH и
отличаются строением радикала R. Формулы и тривиальные названия важнейших
аминокислот приведены в таблице. Для биологического функционирования
аминокислот в составе белков определяющим является полярность радикала R. По
этому признаку аминокислоты разделяют на следующие основные группы (см.
таблицу).
Таблица47.1. Важнейшие a
-аминокислоты RCH(NH2)COOH
|
Формула |
Название |
Обозначение |
pI |
|
Аминокислоты, |
|||
|
|
Глицин |
Gly |
5,97 |
|
|
Аланин |
Ala |
6,0 |
|
|
Валин |
Val |
5,96 |
|
|
Лейцин |
Leu |
5,98 |
|
|
Изолейцин |
Ile |
6,02 |
|
|
Фенилаланин |
Phe |
5,48 |
|
|
Триптофан |
Trp |
5,89 |
|
|
Пролин |
Pro |
6,30 |
|
|
Метионин |
Met |
5,74 |
|
|
Цистин |
(Cys)2 |
5,0 |
|
Аминокислоты, |
|||
|
|
Серин |
Ser |
5,68 |
|
|
Треонин |
Thr |
5,60 |
|
|
Гидроксипролин |
Hyp |
5,8 |
|
|
Аспаргин |
Asn |
5,41 |
|
|
Глутамин |
Gln |
5,65 |
|
Аминокислоты, |
|||
|
|
Лизин |
Lys |
9,74 |
|
|
5-Гидроксилизин |
9,15 |
|
|
|
Аргинин |
Arg |
10,76 |
|
|
Гистидин |
His |
7,59 |
|
Аминокислоты, |
|||
|
|
Аспаргиновая |
Asp |
2,77 |
|
|
Глутаминовая |
Gly |
3,22 |
|
|
Тирозин |
Tyr |
5,66 |
|
|
Цистеин |
Cys |
5,07 |























Аминокислоты, содержащие неполярный радикал R. Такие группы располагаются
внутри молекулы белка и обуславливают гидрофобные взаимодействия.
Аминокислоты, содержащие полярный неионогенный радикал R. Аминокислоты
этого типа имеют в составе бокового радикала полярные группы, не
способные к ионизации в водной среде (спиртовый гидроксил, амидная группа).
Такие группы могут располагаться как внутри, так и на поверхности молекулы
белка. Они участвуют в образовании водородных связей с другими полярными
группами.
Аминокислоты, содержащие радикал R, способный к ионизации в водной среде
с образованием положительно или отрицательно заряженных групп. Такие
аминокислоты содержат в боковом радикале дополнительный основный или кислотный
центр, который в водном растворе может соответственно присоединять или отдавать
протон.

В белках ионогенные группы этих аминокислот располагаются, как правило, на
поверхности молекулы и обуславливают электростатические взаимодействия.
3.2. Стереоизомерия.
Все природные a-аминокислоты (кроме
глицина) являются хиральными соединениями. По конфигурации хирального центра в
положении 2 аминокислоты относят D- или L-ряду.

Природные аминокислоты относятся к L-ряду.
Большинство аминокислот содержат один хиральный центр и имеют два
стереоизомера. Аминокислоты изолейцин, треонин, гидроксипролин, 5-гидроксилизин
и цистин содержат два хиральных центра и имеют (кроме цистина) 4 стереоизомера,
из которых только один встречается в составе белков.
Так, из 4-х стереоизомеров треонина в природе встречается только
(2S,3R)-2-амино-3-гидроксибутановая кислота.

Использование для построения белков только одного вида стереоизомеров имеет
важное значение для формирования их пространственной структуры и обеспечения
биологической активности.
a -Аминокислоты, полученные синтетическим
путем, представляют рацемические смеси, которые необходимо разделять. Наиболее
предпочтительным является ферментативный способ разделения с помощью ферментов
ацилаз, способных гидролизовать N-ацетильные производные только L-a -аминокислот. Ферментативное расщепление
проводят по следующей схеме.
Сначала рацемическую аминокислоту ацилируют уксусным ангидридом:

Затем рацемическую смесь ацетильных производных подвергают ферментативной
обработке. При этом гидролизуется ацетильное производное только L-аминокислоты:

Полученная после ферментативного смесь легко разделяется, так как свободная
L-аминокислота растворяется и в кислотах, и в щелочах, а ацилированная – только
в щелочах.
3.3. Кислотно-основные свойства.
По кислотно-основным свойствам аминокислоты разделяют на три группы.
Нейтральные аминокислоты не содержат в радикале R дополнительных
кислотных или основных центров, способных к ионизации в водной среде. В кислой
среде они существуют в виде однозарядного катиона и являются двухосновными
кислотами по Бренстеду. Как видно на примере аланина, изоэлектрическая точка у
нейтральных аминокислот не равна 7, а лежит в интервале 5,5 – 6,3.

pI=1/2(2,34+9,69)=6,01
Основные аминокислоты содержат в радикале R дополнительный основный
центр. К ним относятся лизин, гистидин и аргинин. В кислой среде они существуют
в виде дикатиона и являются трехосновными кислотами. Изоэлектрическая точка
основных аминокислот, как видно на примере лизина, лежит в области рН выше 7.

pI= 1/2(9,0+10,05)=9,74
Кислые аминокислоты содержат в радикале R дополнительный кислотный
центр. К ним относятся аспаргиновая и глутаминовая кислоты. В кислой среде они
существуют в виде катиона и являются трехосновными кислотами. Изоэлектрическая
точка этих аминокислот лежит в области рН много ниже 7.

pI= 1/2(2,09+3,86)=2,77
Тирозин и цистеин содержат в боковых радикалах слабые кислотные центры,
способные к ионизации при высоких значениях рН.
Важное значение имеет тот факт, что при физиологическом значении рН (~7) ни
одна аминокислота не находится в изоэлектрической точке. В организме все
аминокислоты ионизированы, что обеспечивает им хорошую растворимость в воде.
Различие в кислотно-основных свойствах используется для разделения
аминокислот методом электрофореза и ионообменной хроматографии. При данном
значении рН разные аминокислоты могут иметь разный по величине и знаку
электрический заряд. Например, при рН6 лизин имеет заряд +1 и движется к
катоду, аспаргиновая кислота имеет заряд –1 и перемещается к аноду, а аланин
находится в изоэлектрической точке и не перемещается в электрическом поле.
Таким образом при рН6 они могут быть разделены с помощью электрофореза.
Для разделения аминокислот методом ионообменной хроматографии используют
катионообменные смолы (сульфированный полистирол). Процесс ведут в кислой
среде, когда аминокислоты находятся катионной форме.

Скорость продвижения аминокислот по хроматографической колонке зависит от
силы их электростатических и гидрофобных взаимодействий со смолой. Наиболее
прочно связываются со смолой основные аминокислоты, имеющие наибольший
положительный заряд, наименее прочно – кислые аминокислоты. Наибольшим
гидрофобным связыванием со смолой обладают аминокислоты с неполярными боковыми
радикалами, особенно ароматическими. Таким образом, порядок элюирования
аминокислот следующий. Легче других элюируются кислые аминокислоты (Asp и Glu),
следом за ними идут аминокислоты, содержащие полярные неионогенные группы (Ser,
Thr, Asn, Gln), затем из колонки вымываются аминокислоты с неполярными боковыми
радикалами (Phe, Trp, Ile и др.) и в последнюю очередь элюируются основные аминокислоты
(His, Lys, Arg).
4. Пептиды
Петиды – это полиамиды, построенные из a
-аминокислот. По числу аминокислотных остатков в молекуле пептида различают дипептиды,
трипептиды, тетрапептиды и т.д. Пептиды, содержащие до 10 аминокислотных
остатков, называют олигопептидами, более 10 аминокислотных остатков – полипептидами.
Природные полипептиды, включающие более 100 аминокислотных остатков, называют белками.
4.1. Строение пептидов
Формально пептиды можно рассматривать как продукты поликонденсации аминокислот.

Аминокислотные остатки в пептиде связаны амидными (пептидными)
связями. Один конец цепи, на котором находится аминокислота со свободной
аминогруппой, называют N-концом. Другой конец, на котором находится
аминокислота со свободной карбоксильной группой, называют С-концом. Пептиды
принято записывать и называть, начиная с N-конца.
Название пептида строят на основе тривиальных названий, входящих в его
состав аминокислотных остатков, которые перечисляют, начиная с N-конца. При
этом в названиях всех аминокислот за исключением С-концевой суффикс “ин”
заменяют на суффикс “ил”. Для сокращенного обозначения пептидов используют
трехбуквенные обозначения входящих в его состав аминокислот.

Пептид характеризуется аминокислотным составом и аминокислотной
последовательностью.
Аминокислотный состав пептида может быть установлен путем полного гидролиза
пептида (расщепления до аминокислот) с последующим качественным и
количественным анализом образовавшихся аминокислот методом ионобменной
хроматографии или ГЖХ-анализом сложных эфиров аминокислот. Полный гидролиз
пептидов проводят в кислой среде при кипячении их с 6н. HCl.
Одному и тому же аминокислотному составу отвечает несколько пептидов. Так,
из 2-х разных аминокислот может быть построено 2 дипептида, из трех разных
аминокислот – 6 трипептидов, из n разных аминокислот n! пептидов одинакового
состава. Например, составу Gly:Ala:Val=1:1:1 отвечают следующие 6 трипептидов.
Gly-Ala-Val Gly- Val-Ala Val-Gly-Ala Val-Ala-Gly Ala-Gly-Val Ala-Val-Glu
Таким образом, для полной характеристики пептида необходимо знать его
аминокислотный состав и аминокислотную последовательность.
4.2. Определение аминокислотной последовательности
Для определения аминокислотной последовательности используют комбинацию двух
методов: определение концевых аминокислот и частичный гидролиз.
Определение N-концевых аминокислот.
Метод Сегнера. Пептид обрабатывают 2,4-динитрофтробензолом (ДНФБ), а
затем полностью гидролизуют. Из гидролизата выделяют и идентифицируют
ДНФ-производное N-концевой аминокислоты.

Метод Эдмана состоит во взаимодействии N-концевой аминокислоты с
фенилизотиоцианатом в щелочной среде. При дальнейшей обработке слабой кислотой
без нагревания происходит отщепление от цепи “меченой” концевой аминокислоты в
виде фенилгидантоинового (ФТГ) производного.

Преимущество этого метода состоит в том, что при отщеплении N-концевой
аминокислоты пептид не разрушается и операцию по отщеплению можно повторять.
Метод Эдмана используют в автоматическом приборе – секвенаторе, с помощью
которого можно осуществить 40 – 50 стадий отщепления, идентифицируя полученные
на каждой стадии ФТГ-производные методом газожидкостной хроматографии.
Частичный гидролиз полипептидов
При частичном гидролизе пептиды расщепляются с образованием более коротких
цепей. Частичный гидролиз проводят с помощью ферментов, которые гидролизуют
пептидные связи избирательно, например, только с N-конца (аминопептидазы)
или только с С-конца (карбоксипептидазы). Существуют ферменты,
расщепляющие пептидные связи только между определенными аминокислотами. Меняя
условия гидролиза, можно разбить пептид на различные фрагменты, которые
перекрываются по составляющим их аминокислотным остаткам. Анализ продуктов
частичного гидролиза позволяет воссоздать структуру исходного пептида.
Рассмотрим простейший пример установления структуры трипептида. Частичный
гидролиз по двум разным направлениям трипептида неизвестного строения дает
продукты представленные на схеме.
![]()
Единственный трипептид, структура которого не противоречит продуктам
частичного гидролиза – Gly-Ala-Phe.
Установление аминокислотной последовательности пептидов, содержащих
несколько десятков аминокислотных остатков, – более сложная задача, которая
требует комбинации различных методов.
4.3. Синтез петидов
Синтез пептида с заданной аминокислотной последовательностью – чрезвычайно
сложная задача. В простейшем случае синтеза дипептида из 2-х разных аминокислот
возможно образование 4-х разных продуктов.
![]()
В настоящее время разработана стратегия синтеза пептидов, основанная на
использовании методов активации и защиты функциональных групп на
соответствующих этапах синтеза. Процесс синтеза дипептида включает следующие
стадии:
- защита аминогруппы
N-концевой аминокислоты; - активация карбоксильной
группы N-концевой аминокислоты; - конденсация
модифицированных аминокислот - снятие защитных групп

Таким образом, последовательно присоединяя аминокислоты, шаг за шагом
наращивают цепь полипептида. Такой синтез очень длителен, трудоемок и дает
низкий выход конечного продукта. Основные потери связаны с необходимостью
выделения и очистки продуктов на каждой стадии.
Этих недостатков лишен используемый в настоящее время твердофазный синтез
пептидов. На первой стадии защищенная по аминогруппе С-концевая
аминокислота закрепляется на твердом полимерном носителе (полистироле,
модифицированном введением групп –CH2Cl). После снятия защиты
проводят ацилирование аминогруппы закрепленной на носителе аминокислоты другой
аминокислотой, которая содержит активированную карбоксильную и защищенную
аминогруппу. После снятия защиты проводят следующую стадию ацилирования.
Отмывание продукта от примесей проводят прямо на носителе и лишь после
окончания синтеза полипептид снимают с носителя действием бромистоводородной
кислоты. Твердофазный синтез автоматизирован и проводится с помощью приборов –
автоматических синтезаторов.

Методом твердофазного синтеза получено большое количество пептидов,
содержащих 50 и более аминокислотных остатков, в том числе инсулин (51
аминокислотный остаток) и рибонуклеаза (124 аминокислотных остатка).
