Генетическое расстояние (Genetic distance, GD) — мера генетического различия (дивергенции) между видами, подвидами, или популяциями одного вида. Малое генетическое расстояние означает генетическое сходство, большее генетическое расстояние означает меньшее генетическое сходство.
Существует много параметров, используемых для измерения генетического расстояния.
В простейшем случае генетическое расстояние между двумя популяциями одного вида может быть определено как разница в частотах определённого признака. Например, частота резус-отрицательных людей составляет 50,4 % среди басков, 41,2 % во Франции и 41,1 % в Великобритании. Генетическое расстояние между басками и французами составляет 9,2 %, генетическое расстояние между французами и британцами составляет 0,1 % для признака резус-отрицательный. Генетическое расстояние по нескольким признакам может быть усредненно для вычисления полного генетического расстояния.
Меры генетического расстояния[править | править код]
Существует несколько способов измерения генетического расстояния. Среди них стоит выделить следующие:
Индекс фиксации[править | править код]
Это общепринятый метод определения генетического расстояния. Он принимает минимальное значение 0 и максимальное 1. Значение 0 говорит о генетической идентичности, а величина 1 говорит о том, что две популяции являются разными видами.
Стандартное генетическое расстояние Нэя[править | править код]
Этот метод определения генетического расстояния основан на предположении того, что генетическая разница возникает за счет мутаций и генетического дрейфа. Метод предложил Масатоси Нэи в 1972 году.
Согласно неодарвинистской концепции новый вид возникает в результате дифференциации любой популяции, относящейся к нему; точнее говоря, происходит постепенная дивергенция на уровне генофонда в этой популяции и вида в целом. В основе этой дивергенции лежит постепенная замена одних аллелей определённых генов на другие. В процессе дивергенции двух популяций все меньшее количество генов имеет аллели, которые встречаются в обоих популяциях. Когда заканчивается процесс видообразования, для всех генов существуют аллели, характерные только для одной из популяций. При этом генетическое сходство становится равным нулю и новый вид с морфологической точки зрения становится отличным от первоначального.[источник не указан 4367 дней]
На практике для расчетов генетического расстояния определяют частоту встречаемости в каждой популяции различных форм ряда белков (при этом необходимо, чтобы количество исследуемых белков было достаточным). Число этих форм для каждого определённого белка соответствует числу аллелей гена, кодирующего данный белок. Расчет производится следующим образом. Представим себе, что для какого-то гена G существует i различных аллелей (1, 2, 3 и т. д.). В популяции А эти аллели встречаются с частотой а1, а2, а3 … В популяции В эта частота соответствует b1, b2, b3 … Исходя из этого, можно вычислить степень генетического сходства данных популяций по данному гену:

Это уравнение позволяет вычислить нормализованную вероятность того, что два аллеля из разных популяций будут идентичными.
Если провести подобные расчеты для n генов, принадлежащих обоим популяциям, то можно будет вычислить среднюю арифметическую величину 

Можно также рассчитать величины 




Генетическое расстояние (D) вычисляется по следующей формуле:

Значения генетического сходства могут варьировать от 0 (между популяциями нет общих аллелей), до 1 (частоты аллелей в популяции одинаковые).
Если индекс генетического сходства между двумя выборками равен 0.65, то можно сказать что 65 % аллелей между выборками являются общими.
Если генетическое расстояние составляет 0,62 (именно такое расстояние между человеком и шимпанзе), то из 100 генов, принадлежащих генофонду обоих видов, 62 (то есть примерно 2 / 3) имеют аллели, принадлежащие только одному из них. Если же генетическое расстояние между двумя популяциями составляет 1, то 100 % генов этих популяций имеют аллели, принадлежащие только к одному из них. Такое расстояние соответствует завершению постепенной дивергенции популяции исходного вида и окончательному отделению нового вида. Степень генетического различия между двумя подвидами, относящихся к одному виду, соответствует генетическому расстоянию, равному 0,17−0,22. Генетическое расстояние между двумя видами-двойниками (то есть популяциями, которые не отличаются по морфологическим признакам, но лишаются способности к скрещиванию) составляет 0,50-0,60.
Индекс Кавалли-Сфорца и Эдвардса (1967)[править | править код]
Этот метод определения генетического расстояния основан на предположении того, что генетическая разница возникает только за счет генетического дрейфа
Генетическое расстояние в области экономики языка[править | править код]
Экономисты Энрико Сполаоре и Ромен Вациарг выдвигают гипотезу о том, что существует значительная положительная взаимосвязь между длительным родством популяций (которое измеряется таким показателем, как генетическое расстояние) и рядом показателей культурных различий[1]. Для этого авторы рассматривают простую модель, связывающую генетическую дистанцию с культурной дистанцией.
Исследование культурной дистанции основывается на трёх измерениях:
- языковой дистанции;
- религиозной дистанции;
- ценностях, культурных нормах, убеждениях.
Э. Сполаоре и Р. Вациарг используют следующую формулу для вычисления генетической дистанции: 







Если допустить, что 


Формула для вычисления вероятности, что две случайно выбранные аллели гетерозиготны в популяции 

Средняя частота генов аллели 1 в двух популяциях определяется следующим образом: 

Уравнение для вычисления гетерозиготности в двух популяциях будет описываться следующим образом: 




В общей сложности, если две популяции имеют одинаковые частоты аллелей (




Генетическое расстояние и время отделения популяций[править | править код]
Время отделения популяций — время, которое прошло с момента, когда две популяции имели последних общих предков. Его формулу можно записать следующим образом: 


При небольшом значении 

Это означает, что генетическое расстояние между двумя двоюродными популяциями примерно пропорционально времени, прошедшему с тех пор, как предки двух популяций отделились и образовали отдельные популяции. Таким образом, в этом отношении можно интерпретировать генетическое расстояние как показатель времени, прошедшего с тех пор, как две популяции имели общего предка.
Измерения культурной дистанции[править | править код]
Языковая дистанция[править | править код]
Для измерения языковой дистанции используют, как правило, два метода: языковые деревья и лексикостатистику.
Измерение по методу языковых деревьев заключается в том, что языки группируются на основе предполагаемого сходства между ними. Например, испанский имеет следующую классификацию: индоевропейская семья — италийский язык — романский язык — западно-романский язык — иберо-романский язык — западно-иберийский язык — испанский язык; а румынский язык, в свою очередь, будет классифицирован следующим образом: индоевропейская семья — италийский язык — романский язык — балкано-романский язык — румынский язык. Из этого следует, что оба языка имеют три общих узла. Так, при изменении количества общих узлов соответственно будет изменяться лингвистическое расстояние между языками.
Для определения количества общих узлов между двумя языками применяется следующая формула: 







Языковая дистанция по методу языковых деревьев (


Вычисление языкового расстояния на основе метода лексикостатистики заключается в том, что рассматриваются слова из разных языков, передающие некоторые общие значения (из списка Сводеша), и исследуется, являются ли эти слова родственными и происходят ли они от одного языка-предка. Например, английское слово ‘swim’ и немецкое ‘schwimmen’ происходят из староанглийского слова ‘swimman’. Следовательно, эти слова являются родственными.
Таким же образом, как и при вычислении 

Религиозная дистанция[править | править код]
Религиозная дистанция высчитывается аналогично вычислению языкового расстояния по методу языковых деревьев и описывает взаимосвязь между мировыми религиями. Соответственно, количество общих узлов между религиями является показателем религиозной близости.
Ценности, культурные нормы и убеждения[править | править код]
В качестве показателей культурных норм, ценностей и установок респондента могут быть использованы ответы на вопросы социальных опросов. Если провести аналогию с генетикой, то можно сказать, что вопросы соответствуют локусам генов, в то время как конкретные ответы будут соотнесены с аллелями. Таким образом, различия между группами населения в ответах на конкретный вопрос могут быть использованы для расчета культурной дистанции между странами по этому конкретному вопросу.
Сложности, которые могут возникнуть при вычислении данных культурных индексов:
- выбор вопросов (для получения репрезентативных результатов, вместо произвольного выбора вопросов необходимо рассматривать набор всех вопросов, связанных с ценностями, которые фигурируют в интегрированном вопроснике World Values Survey[2] 1981—2010, которые перечислены WVS как категории от A до G. Однако при расчете сводных индексов культурной дистанции, которые суммируются по вопросам, важно иметь одинаковое количество вопросов для каждой пары стран);
- выбор функциональной формы для вычисления расстояний для каждого вопроса. Так, для бинарных вопросов вычисление культурного расстояния
между странами 1 и 2 будет проводиться по следующей формуле:
. Для небинарных вопросов будет использоваться следующее уравнение:
.
- суммирование расстояния по конкретным вопросам для получения сводных показателей культурной дистанции.
Родство и культура: простая концептуальная основа[править | править код]
Предки передают большое количество своих черт потомкам не только биологически, но и культурно. Популяции, которые более тесно связаны между собой, будут иметь меньше времени, чтобы отделиться друг от друга в наборе таких культурных черт как язык, религия, традиции, привычки и ценности. Этот процесс устанавливает тесную связь между родословной,
измеряется генетической дистанцией и чертами, передающимися в культуре: генетическая дистанция и меметическая дистанция должны иметь положительную корреляцию. Стилизованная и формальная модель, адаптированная из Spolaore и Wacziarg (2009, 2012) иллюстрирует эти отношения в упрощенном виде.
Возьмем три популяции: i = 1, 2, 3 живущих в настоящем. Популяции 1 и 2 происходят от общего последнего населения предков. Популяция 3 имеет несколько общих предков с популяциями 1 и 2, идущими дальше во времени. Тем самым, популяция 3 менее близка к популяциям 1 и 2, которые можно ассоциировать с родными братом и сестрой, в то время как популяция 3 является дальним двоюродным родственником.
Говоря о результатах в разделе 6.2.2 «Генетическая дистанция между человеческими популяциями», можем предположить генетическое расстояние dg (i, j) между популяцией i и популяцией j как время, в которое они были одной популяцией. Заключаем, что генетическое расстояние dg (1, 2) между популяцией 1 и популяцией 2 меньше, чем генетическое расстояние между популяцией 1 и популяцией 3, а также меньше чем генетическое расстояние между популяцией 2 и популяцией 3:
dg (1, 2) = F < dg (1, 3) = dg (2, 3) = F
Насколько далеки друг от друга по культурным признакам эти популяции? Культурное расхождение зависит от сложных процессов передачи от поколения к поколению большого набора культурных черт, полезно фокусироваться на простейшем возможном механизме культурной передачи с вариацией, тогда как культура фиксируется только одной чертой (или мемом), которая может изображаться точкой на прямой. За каждый период t популяция i обладает культурными чертами ci(t), которые с вариациями унаследованы от популяции-предка,
который имел черты ci (t — 1) согласно:
ci(t) = ci(t − 1) + εi(t)
Можно полагать, что простейший возможный механизм для вариации: культурная смена как случайная прогулка.
В заключение, большее генетическое расстояние связано с большей дистанцией в культурных чертах. Это отношение не детерминировано. Некоторые пары популяций, которые являются более дальними родственниками, могут иметь более сходные культурные особенности.
черты, чем две более близкородственные популяции, но такой исход менее вероятен.
Генетическая дистанция и дистанция в культурно передаваемых чертах, таких как язык, религия, ценности ожидаются в положительной корреляции.
Родство и культура: эмпирические данные[править | править код]
Эмпирическое исследование взаимосвязи между генетическими и меметическими расстояниями проводится с целью проверки гипотезы о том, что более длительное время разделения на самом деле положительно связано с различиями в языке, религии и нормах, ценностях и взглядах.
Генетическая дистанция и лингвистическая дистанция[править | править код]
Показатели лингвистической и генетической дистанции должны иметь положительную корреляцию. Кавалли-Сфорца проследил маленькую генетическую связь между языковыми группами. Филогенетические деревья и лингвистические деревья имеют тенденцию
быть похожими друг на друга. Идеальных отношений не стоит ждать по нескольким причинам:
Во-первых, лингвистические данные основанные на деревьях имеют дискретное количество узлов, тогда как генетическое расстояние, основанное на большом количестве аллелей, как в случае используемого нами индекса, равно
непрерывная мера времени разделения.
Во-вторых, функциональные формы для
меры генетической дистанции (FST) и лингвистической дистанции различны.
В-третьих, успешные группы, завоевывающие территории отдельных языковых групп, могут навязывать свой язык не навязывая свои гены. Так было, например, с мадьярским завоеванием Венгрии: образовавшийся в результате язык принадлежал уральской семье, но генетическая примесь мадьяр была настолько ограничена, что венгры генетически очень близки к другим славянским популяциям, например к полякам. Другим ярким примером служат движения населения, последовавшие за открытие Нового Света, в частности работорговля: нынешние потомки бывших рабов не говорят на исконных западноафриканских языках своих предков. Точно так же нынешние жители Соединенных Штатов преимущественно говорят по-английски, в то время как их предки происходили из разных языковых групп. Таким образом, современные миграции послужили разрыву связи между генетическими и языковая дистанция.
В таблице 6.2 на панели А исследуются основные корреляции. Мы обнаруживаем, что наши различные меры языковой дистанции сильно коррелируют между собой.
В таблице 6.3 представлены регрессии наших различных показателей языковой дистанции, генетической дистанции, с контролем или без контроля широкого спектра показателей, географическое разделение — в том числе геодезическое расстояние, абсолютная разница в долготе и широте и т. д.
Генетическая дистанция и религиозная дистанция[править | править код]
Религиозные верования, также как и язык передаются из поколения в поколение, что позволяет ожидать положительной корреляции между религиозной дистанцией и генетическим расстоянием.
Несколько факторов, которые ограничивают степень корреляции между религиозной и генеалогической дистанциями:
- Они могут найти свой источник древних религиозных верований. Несколько крупных мировых религий появились относительно недавно (иудаизм).
- Генетически сходные популяции часто исповедуют разные религиозные верования. Христианство и ислам произошли среди близкородственных популяций на Ближнем Востоке.
- Религиозный верования передаются по горизонтали через завоевания и преобразования, так как легче изменить религию, чем язык.
- Вышеупомянутые различия функциональной формы между показателями языковой и генетической дистанции с той же силой применимы к показателям религиозного расстояния.
Религиозная дистанция положительно коррелирует с генетической дистанцией. Первое доказательство представлено на Панели B. Таблицы 6.2. Мы видим, например, взвешенную религиозную дистанцию, основанную на религиозном дереве мехам, ферон и лаитин (F — RD) имеет корреляцию 0,18 со взвешенным генетическим расстоянием.
В таблицах 6.5 и 6.6 представлены регрессионные данные, опять же с учётом или без учёта географического расстояния для каждого из четырёх показателей религиозного расстояния. Во всех спецификациях, кроме одной, генетическая дистанция дает положительный результат.
Генетическая дистанция и культурная дистанция[править | править код]
Последнее исследование касается взаимосвязи между генетической дистанцией и дистанция в нормах, ценностях и установках. При нулевой гипотезе об отсутствии связи между генетическими и культурными расстояниями мы ожидаем, что 5 %
корреляции должны быть значимыми (2,5 % положительных и значимых) и распределение корреляций должно быть сосредоточено вокруг нуля. Таблица 6.5 представляет
гистограмма выборочных корреляций между двусторонним расстоянием для каждого вопроса, и взвешенное генетическое расстояние для полного набора из 740 вопросов.
Хотя эти результаты информативны, они объединяют вопросы по очень разным предметам и разным типам (бинарные и небинарные). В таблице 6.7 представлены простые корреляции. Генетическое расстояние имеет корреляцию
0,27 с нашей суммарной мерой культурной дистанции.
В таблице 6.9 вопросы разбиты по категориям. На нижней панели с географическим контролем мы видим положительные и значительные эффекты. Наибольшее влияние в количественном отношении имеют категории A (восприятие жизни), E (политика и общество) и F (религия и мораль). Будущая работа должна быть направлена на углубление более глубоко в характеристиках вопросов, наиболее тесно связанных с родовым расстоянием.
Выводы[править | править код]
Генетическая дистанция — это суммарная мера различий в широком спектре человеческих черт, передаваемых из поколения в поколение. Мы сосредоточились на языке, религии и ценностей, находя эмпирические доказательства положительной корреляции между генетической дистанцией и языковой, религиозной и культурной дистанцией. Важно отметить, что генетическое расстояние не сильно коррелирует только с небольшим и специфическим подмножеством различий в культурных особенностях. Наоборот, генетическая дистанция, как правило, широко и значимо коррелирует с широким диапазоном культурных различий черты.
Таким образом, в то время как специфические корреляции с отдельными наборами признаков обычно умеренны по величине, существует общая связь между происхождением и культуры, в соответствии с концептуальной структурой, в которой широкий спектр культурных черт передается с вариациями от поколения к поколению с течением времени. Генетическое расстояние является полезной сводной мерой, фиксирующей различия в этом широком диапазоне культурных особенностей.
См. также[править | править код]
- Коэффициент инбридинга
Примечания[править | править код]
- ↑ E. Spolaore, R. Wacziarg. Ancestry, Language and Culture (англ.) // The Palgrave Handbook of Economics and Language. — 2015. — P. 174—210.
- ↑ World Values Survey (англ.). Дата обращения: 24 марта 2022. Архивировано 25 марта 2022 года.
Ссылки[править | править код]
- Генетическое расстояние на сайте Genopus.ru (недоступная ссылка) (рус.)
- A review of mutation processes and methods of phylogenetic inference, David B. Goldstein, David D. Pollock
- Stanford University website on genetic distance
- The Estimation of Genetic Distance and Population Substructure from Microsatellite allele frequency data., Brent W. Murray (May 1996), McMaster University website on genetic distance
- Computing distance by stepwise genetic distance model, web pages of Bruce Walsh at the Department of Ecology and Evolutionary Biology at the University of Arizona
- Britten RJ. Divergence between samples of chimpanzee and human DNA sequences is 5 %, counting indels. PNAS; 2002:13633
Сцепленное наследование генов
Необходимо запомнить
ВАЖНО!
Сцепление генов – это совместное наследование генов, расположенных в одной и той же хромосоме. Количество групп сцепления соответствует гаплоидному числу хромосом, то есть у дрозофилы 4. Природу сцепленного наследования объяснил Морган с сотрудниками. В качестве объекта исследования они избрали плодовую муху дрозофилу, которая оказалась очень удобной моделью для изучения данного феномена, так в клетках её тела находится только 4 пары хромосом и имеет место высокая скорость размножения (в течение года можно исследовать более 20-ти поколений). Итак, сцепленными признаками называются признаки, которые контролируются генами, расположенными в одной хромосоме. Естественно, что они передаются вместе в случаях полного сцепления (закон Моргана). Полное сцепление встречается редко, обычно – неполное, из-за влияния кроссинговера (перекрещивания и обмена участками гомологичных хромосом в процессе мейоза). То есть, гены одной хромосомы переходят в другую, гомологичную ей.
Частота кроссинговера зависит от расстояния между генами. Чем ближе друг к другу расположены гены в хромосоме, тем сильнее между ними сцепление и тем реже происходит их расхождение при кроссинговере, и, наоборот, чем дальше друг от друга отстоят гены, тем слабее сцепление между ними и тем чаще возможно его нарушение.

Рис. 1
На рисунке 1 слева: расстояние между генами А и В маленькое, вероятность разрыва хроматиды именно между А и В невелика, поэтому сцепление полное, хромосомы в гаметах идентичны родительским (два типа), других вариантов не появляется.
На рисунке 1 справа: расстояние между генами А и В большое, повышается вероятность разрыва хроматиды и последующего воссоединения крест-накрест именно между А и В, поэтому сцепление неполное, хромосомы в гаметах образуются четырёх типов – 2 идентичные родительским (некроссоверные) + 2 кроссоверных варианта.
Количество разных типов гамет будет зависеть от частоты кроссинговера или расстояния между анализируемыми генами. Расстояние между генами исчисляется в морганидах – единицах расстояния между генами, находящимися в одной хромосоме. 1 морганида соответствует 1 % кроссинговера. Такая зависимость между расстояниями и частотой кроссинговера прослеживается только до 50 морганид. Частота кроссинговера между определенной парой генов – довольно постоянная величина (хотя радиация, химические вещества, гормоны, лекарства влияют на нее; например, высокая температура стимулирует кроссинговер).
Пример, основанный на опытах Моргана

Рис. 2
Гены, расположенные в одной хромосоме, наследуются совместно. Фенотипы А – серое тело, нормальные крылья (повторяет материнскую форму). Б – тёмное тело, короткие крылья (повторяет отцовскую форму). В – серое тело, короткие крылья (отличается от родителей). Г – тёмное тело, нормальные крылья (отличается от родителей). В и Г получены в результате кроссинговера в мейозе.
Если скрестить мушку дрозофилу, имеющую серое тело и нормальные крылья (на рисунке самка), с мушкой, обладающей тёмной окраской и зачаточными (короткими) крыльями (на рисунке самец), то в первом поколении гибридов все мухи будут серыми с нормальными крыльями (А). Это гетерозиготы по двум парам аллельных генов, причём ген, определяющий серую окраску брюшка, доминирует над тёмной окраской, а ген, обусловливающий развитие нормальных крыльев, – доминирует над геном недоразвитых крыльев.
При анализирующем скрещивании гибрида F1 с гомозиготной рецессивной дрозофилой (Б) подавляющее большинство потомков F2 будет сходно с родительскими формами.
Это происходит потому, что гены, отвечающие за серое тело и нормальные крылья– сцепленные гены, также как и гены, отвечающие за тёмное тело и короткие крылья, т. е. они находятся в одной хромосоме. наследование сцепленных генов называют сцепленным наследованием.
Сцепление может нарушаться. Это доказывает наличие особей В и Г на рисунке, т. е. если бы сцепление не нарушалось, то этих особей бы не существовало, однако они есть. Это происходит в результате кроссинговера, который и нарушает сцепленность этих генов.

Рис. 3
На рисунке 3 опыт Моргана отображен подробно. Несцепленное наследование: два гена находятся в разных хромосомах, гетерозигота с равной вероятностью дает четыре типа гамет:

Рис. 4
Сцепленное наследование: два гена находятся в одной хромосоме.
а) При полном сцеплении гетерозигота дает только два типа гамет.
б) При неполном сцеплении гетрозигота дает четыре типа гамет, но не с равной вероятностью.

Рис. 5
На вышесказанном строится хромосомная теория наследственности Моргана:
1. Гены находятся в хромосомах и расположены в линейной последовательности на определенных расстояниях друг от друга.
2. Гены, расположенные в одной хромосоме, составляют группу сцепления. Число групп сцепления равно гаплоидному числу хромосом. Признаки, гены которых находятся в одной хромосоме, наследуются сцепленно (т. е. в тех же сочетаниях, в которых они были в хромосомах исходных родительских форм).
3. Новые сочетания генов, расположенных в одной паре хромосом, могут возникать в результате кроссинговера в процессе мейоза. Частота кроссинговера зависит от расстояния между генами.
4. Учитывая линейное расположение генов в хромосоме и частоту кроссинговера как показателя расстояния между генами, можно построить карты хромосом. За единицу расстояния между генами принята частота кроссинговера равная 1 % (морганида, сантиморган, сМ).
Задачи на нахождение расстояния между генами
Решение задачи на определение вероятности рождения здоровый и больных детей
Сцепленные гены — это неаллельные гены, которые находятся на одной хромосоме. Гены, которые расположены на одной хромосоме, называются группой сцепления.
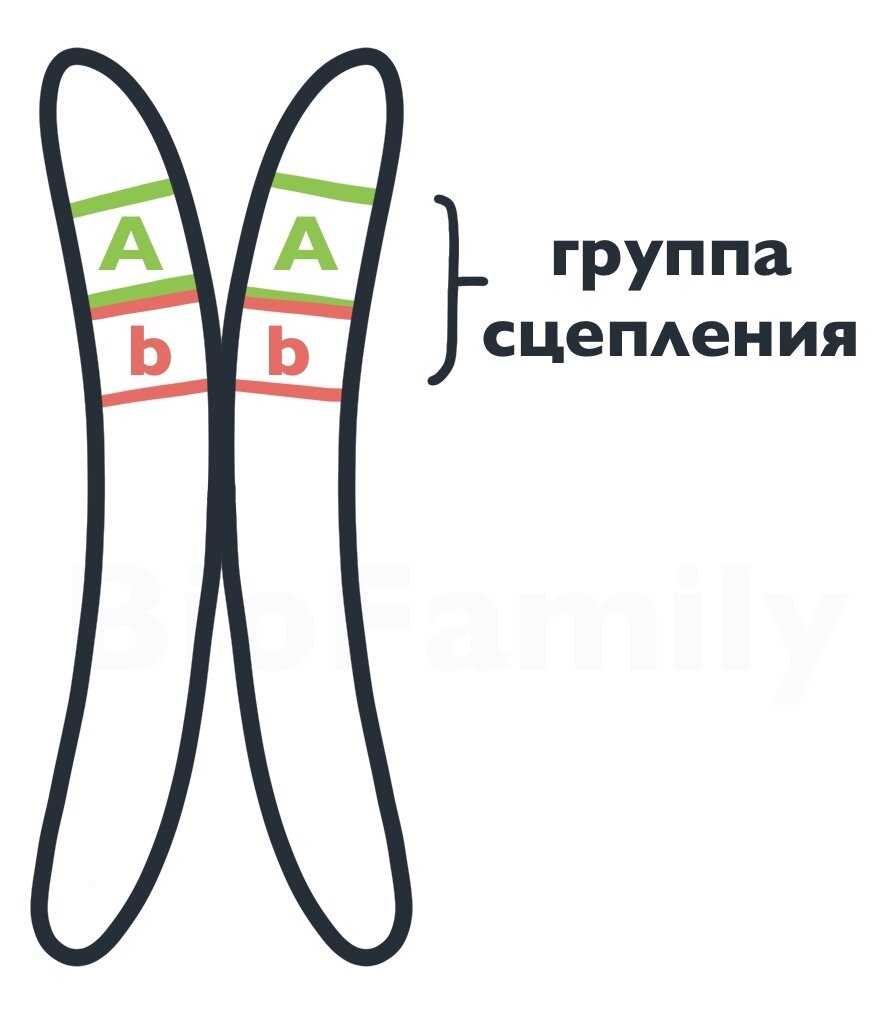
Различают полное сцепление генов и неполное сцепление (нарушение сцепления).
Полное сцепление генов
При кроссинговере (обменом участками гомологичных хромосом) хромосомы обмениваются не конкретными генами, а большими участками. Из-за этого гены, расположенные рядом, не могут разойтись по разным хромосомам. Так возникает сцепленное наследование, не нарушенное кроссинговером (полное сцепление).
Приведём пример задачи на сцепленное наследование генов…
Текст задания:
У томатов гены, определяющие высокий рост стебля и шаровидную форму плодов, сцеплены и доминируют над генами, определяющими карликовость и грушевидную форму плодов. Какое потомство следует ожидать от скрещивания гетерозиготного по обоим признакам растения с карликовым, имеющим плоды грушевидной формы?
Решение:
Пусть высокий рост будет обозначаться «А», а карликовость — «а». Шаровидная форма тогда будет «В», а грушевидная — «b». В задаче не сказано про промежуточные признаки, поэтому доминирование полное.
Представим, если в задаче не было бы сцепления, и гены располагались в разных парах гомологичных хромосом. Тогда бы генотип AaBb давал бы 4 типа гамет:
А в данной задаче нам говорится, что сцеплены «A» с «B», в то время как «а» сцеплено с «b». Это значит, что гаметы «Ab» и «aB» не образуются.
В результате эта дигетерозигота даст всего 2 типа гамет вместо 4:
Вот как на бумаге выглядит полное решение данной задачи.
В задаче должна быть легенда (то, что нам дано), она пишется справа или слева и заключается в прямоугольник. Необходимо написать скрещивание (Р), гаметы (G) и поколение (F1).
Рядом с каждым генотипом должен быть подписан его фенотип (например, AaBb — высок. шар.) !!!
Текстовое объяснение в задаче следует после, в данном случае оно должно выглядеть примерно так:
1) В потомстве от дигетерозиготы (высокое растение с шаровидными плодами) и рецессивной дигомозиготы (карликовое растение с грушевидными плодами) 50% особей получаются высокими с шаровидными плодами (AaBb), а 50% особей — карликовыми с грушевидными (aabb).
2) Фенотипическое расщепление 1:1.
3) Оно обусловлено попарным сцеплением гена высокого роста с геном шаровидной формы плодов и гена карликовости с геном грушевидной формы плодов.
4) Эти гены расположены на одной хромосоме и образуют группу сцепления. Сцепленный тип наследования генов изучал Морган.
Текстовое объяснение должно быть всегда!!! Старайтесь описывать каждое своё действие в задаче. За присутствие лишних пунктов в объяснении баллы не снижают, а вот за их отсутствие — очень любят.
Нарушение сцепления
Существуют частные случаи, в которых гены расположены в одной хромосоме , но недостаточно близко друг к другу. Из-за этого кроссинговер разъединяет их, но редко.
В случае нарушения сцепления у дигетерозиготной особи образуются 2 типа гамет:
- Гаметы, полученные в результате сцепления (в этом случае сочетание генов в хромосоме будет таким же, как было у родителей)
- Кроссоверные гаметы, полученные в результате расхождения генов (в этом случае возникаю новые варианты сочетания генов на хромосоме, которых не было у родителей)
Гамет, образованных в результате сцепления, всегда больше!
Морганиды — это условная величина, которую применяют для обозначения расстояния между генами на одной хромосоме.
Найти расстояние между генами можно, разделив количество рекомбинантных особей (полученных в результате слияния кроссоверных гамет) на общее число особей. Так, мы находим процент кроссинговера в скрещивании, который равен числу морганид.
Приведём пример задачи на нарушение сцепления генов…
Текст задачи:
При анализирующем скрещивании высокорослого растения с цельной листовой пластинкой получили 9 высокорослых растений с цельной листовой пластинкой, 42 высокорослых растения с расчленённой листовой пластинкой, 40 карликовых растений с цельной листовой пластинкой и 10 карликовых растений с расчлененной листовой пластинкой. Определите генотипы и фенотипы родителей. Определите генотипы потомства. Объясните появление 4 фенотипических групп.
Решение:
Пусть за рост отвечает гены А/а, а за листовую пластинку — B/b.
Нам не сказано, какой признак доминантный, а какой — рецессивный. Но есть подсказка: в результате анализирующего скрещивания (это значит, что организм скрестили с «aabb») получилось 4 фенотипические группы. Это значит, что вторая исходная особь должна быть гетерозиготой (AaBb). Почему? Потому что в другом случае фенотипов было бы 2, либо вообще 1. Проверьте сами.
Если мы знаем, что исходная особь была гетерозиготой, то, зная её фенотип, можно узнать и доминантные признаки. «При анализирующем скрещивании высокорослого растения с цельной листовой пластинкой…» — сделаем выводы:
AA/Aa (можно просто написать A_) — высокий рост.
aa — карликовость.
B_ — цельная листовая пластинка.
bb — расчленённая листовая пластинка.
Составим схему скрещивания…
Из условия мы знаем фенотипы и численность полученного потомства.
Теперь самое важное: зачем нам числа. Числа показывают то, какие особи являются рекомбинантными — полученными в результате слияния кроссоверных гамет, а какие особи являются результатом слияния гамет без кроссинговера (со сцеплением).
Рекомбинантных особей всегда меньше!
Соответственно, числа 9 и 10 подсказывают нам, что генотипы AaBb и aabb — рекомбинантные. А значит другие генотипы: Aabb (высокий расчленённый) и aaBb (карликовый цельный) — результат сцепления.
Таким образом, сцеплен ген высокого роста (А) с геном расчленённой листовой пластинки (b). Ген карликовости (a) сцеплен с геном цельной пластинки (B).
Отвечаем на последний вопрос задачи:
«Преимущественно образовались две фенотипические группы: 42 Aabb (высокие, расчленённые листья) и 40 aaBb (карлики, цельные листья), потому что в гаметах Ab и aB сцеплен ген высокого роста (А) с геном расчленённой листовой пластинки (b), в то время как ген карликовости (a) сцеплен с геном цельной пластинки (B). 9 AaBb (высокие, цельные листья) и 10 aabb (карлики, расчленённые листья) образовались благодаря кроссоверным гаметам AB и ab, которые получились в результате кроссинговера.» Задача решена.
В задании не спрашивается этого, но для наглядности давайте найдём расстояние между данными генами.
Количество рекомбинантных особей: 19. Количество всех особей: 101.
19/101 = 0,188… Соответственно, кроссинговер происходит в 18,8 %. Это и есть расстояние между генами — 18,8 морганид.
Также в задании могут фигурировать не числа, а проценты. Логика выполнения должна быть такая же. Если вы видите четыре фенотипические группы: две малочисленные и две многочисленные, то перед вами скорее всего задача на нарушение сцепления.
Поздравляю с успешным освоением новой темы!
Хочешь узнать, как прошло поступление в этом году? Мы подготовили подборку проходных баллов медицинских вузов (тыкай и забирай полный файл ВКонтакте)
Получить тест по теме в ЛС →
Примерное время выполнения: 50 минут.
Мы используем сервис «РЕШУ ЕГЭ» как самый удобный в коммуникации между учителем и учеником. На сервисе возможна авторизация через ВК.
В этот раз задания будут только из второй части. Так, они не будут проверяться автоматически. Выполнить их следует на листе. Обязательно проверьте их сами — не бойтесь подглядывать в ответы.
При
решении задач предыдущего параграфа
мы исходили из того, что генетические
карты хромосом уже имеются. Это позволяло
нам, путем вычисления, установить
проценты кроссоверных и некроссоверных
гамет. Однако у многих объектов гены не
картированы. В генетике человека
составление генетических карт только
начинается. Поэтому важно ознакомиться
и с задачами по определению расстояния
между генами и составлению генетических
карт хромосом.
Для
решения вопроса, относятся ли два гена
к одной или разным группам сцепления и
для определения расстояния между двумя
генами одной и той же группы сцепления,
ставится опыт дигибридного
анализирующего скрещивания. Подсчет
фенотипов потомства с последующими
простыми вычислениями позволяет
ответить на оба поставленных вопроса.
У человека дело обстоит сложнее. Нужно
выбирать такие родословные, в которых
есть браки, соответствующие требованиям
дигибридного анализирующего скрещивания.
Относительная редкость браков такого
рода затрудняет определение групп
сцепления и расстояний между генами.
У
дрозофилы для анализирующего скрещивания
берут дигибридную по исследуемым генам
самку и скрещивают ее с рецессивным
самцом. Чтобы показать, какие при этом
могут быть получены результаты, разберем
задачи 95 и 96.
По
условиям задачи 95 дигетерозиготная по
генам А и В самка (генотип АаВb) скрещена
с рецессивным по обоим генам самцом
(aabb).
Если гены находятся в разных хромосомах,
то при образовании самкой гамет гены
будут свободно комбинироваться и мы
получим четыре типа гамет (АВ, Аb, аВ, аb)
в равных количествах (по 25% каждого
типа). Самец образует один тип гамет —
ab,
который не будет влиять на фенотип
потомства, так как оба гена в сперматозоидах
рецессивны. Вследствие этого фенотип
потомства будет целиком определяться
генами, находящимися в гаметах самки.
Результаты скрещивания можно представить
в виде схемы (см. схему 40).
Схема
40. Анализирующее дигибридное скрещивание
при свободном комбинировании.
Рассматривая
схему 40, нужно обратить внимание на
следующие два итога. Во-первых, формула
фенотипов первого поколения при
анализирующем скрещивании (АВ, Ab, аВ,
ab) полностью совпадает с формулами гамет
самки. Из этого видно, что анализирующее
скрещивание дает возможность установить
по фенотипу потомства формулы гамет
самок. Во-вторых, при анализирующем
скрещивании все четыре фенотипа потомков
появляются в равных количествах (по
25%). Это характерно для свободного
комбинирования генов, и доказывает,
что гены А и В локализованы в разных
хромосомах (если гены А и В находятся в
одной хромосоме на большом расстоянии
друг от друга, расщепление в первом
поколении получается близким к расщеплению
при свободном комбинировании генов).
Разберем
задачу 96. Дигетерозиготная по генам М
и N самка дрозофилы скрещена с рецессивным
самцом. При этом получено следующее
расщепление по фенотипу:
MN —
47%; Mn
— 3%; mN — 3%; mn — 47%.
Представим данные опыта в виде генетической
схемы (см. схему 41).
Схема
41. Анализирующее дигибридное скрещивание
при сцеплении генов.
Отметим
значительное различие в числе особей
разных фенотипов. Отношение фенотипов
в
F1
составляет в процентах— 47 : 3 : 3 : 47, вместо
ожидаемого при свободном комбинировании
отношения 25 : 25 : 25 : 25. Это свидетельствует
о том, что гены М и N наследуются сцепленно,
то есть находятся в одной и той же
хромосоме. Данные задачи 96 позволяют
ответить еще на два вопроса: 1) как
комбинируются доминантные и рецессивные
аллели в парных хромосомах; 2) на каком
расстоянии они находятся?
В
условиях задачи не указано, как расположены
гены М,
m, N, n
в парных хромосомах самки, но по фенотипу
потомства видно, что гены М и
N,
а также
m
и п обнаруживают высокую степень
сцепления 47+47=94%. Следовательно, в парных
хромосомах самки ген М комбинируется
с N,
а m
с n.
Генотип
самки
Расстояние
между генами М и N определяется по
суммарному проценту рекомбинантов.
Особей Мп, возникших из кроссоверных
гамет 3% и особей
mN,
тоже 3%. Таким образом общее количество
рекомбинантов 6% и расстояние между
генами М и N (а также m и n) равно 6
морганидам.
В
задаче 102 б) рассматривается особый
случай анализирующего скрещивания.
Дигетерозиготная самка скрещивается
с доминантным (а не рецессивным) по обоим
признакам самцом. Скрещивание такого
рода позволяет анализировать генотип
самки только в том случае, если гены
локализованы в Х-хромосоме.
Разберем
задачу. У дрозофилы рецессивный ген
cut
(с) обусловливает вырезки на крыльях, а
рецессивный ген
tan (t)
— смуглее тело. Оба гена локализованы
в Х-хромосоме. Можно ли установить
расстояние между генами
cut,
скрещивая дигетерозиготную самку с
доминантным по обоим генам самцом.
Чтобы ответить на этот вопрос рассмотрим
схему скрещивания (см. схему 42). Для
упрощения рассуждений при
разборе
задачи примем, что самка имеет генотип
Схема
показывает, что все самки первого
поколения имеют оба доминантных признака.
Это вполне понятно, так как все они
получили Х-хромосому с двумя доминантными
генами от отца. Эти гены проявят свое
действие независимо от того, с каким
типом яйцеклеток матери они сольются
при оплодотворении. Другими словами,
фенотип дочерей в этом опыте определяется
отцом. Сыновья в этом опыте получают от
отцд Y-хромосому, не несущую аллелей С,
с и Т, t. Поэтому их фенотип целиком
определяется генами, полученными через
яйцеклетку матери, и их расщепление по
фенотипу будет отражать количество
некроссоверпых и кроссоверных гамет у
матери. Таким образом, при постановке
анализирующего скрещивания дигибридной
самки с доминантным самцом для исследования
сцепления генов, локализованных в
Х-хромосоме, следует учитывать
расщепление только среди сыновей.
Условия
задачи 102 в), в которой сказано, что число
рекомбинантов среди сыновей составляет
3,75%
Ct
и 3,75% сТ, позволяют утверждать, что гены
с и t расположены в Х-хромосоме на
расстоянии 3,75+3,75=7,5 морганид.
Разберем
одну из сложных задач этого параграфа.
В задаче 104 требуется определить
расстояние между двумя генами из
Х-хромосомы человека: геном, обуславливающим
цветовую слепоту (с) и геном мышечной
дистрофии Дюшена (d).
Схема
42. Анализирующее скрещивание дигибридной
самки с доминантным самцом, при локализации
генов в Х-хромосоме.
Согласно
родословной муж и жена здоровы и обладают
нормальным цветовым зрением. Даны
сведения о родителях жены: ее отец
страдал мышечной дистрофией, а мать
нарушением цветового зрения. Даны также
сведения об их потомстве. Из одиннадцати
детей, родившихся в этой семье, 3 дочери
и 1 сын нормальны по обоим признакам, 3
сына страдали мышечной дистрофией,
3 сына — цветовой слепотой, и один сын
— обоими болезнями.
Определим
генотипы родителей. Так как по генам,
локализованным в Х-хромосоме, муж
гемизиготен, то его генотип определяется
по фенотипу. В единственной Х-хромосоме
он имеет гены С и
D.
Чтобы установить генотип жены, нужно
использовать как сведения о детях,
так и сведения о родителях жены.
Сведения о дочерях не могут иметь
значения для определения генотипа
матери, так как дочери получили Х-хромосому
с двумя доминантными генами от отца,
что и определяет их фенотип, независимо
от того, какие гены они получили от
матери. Напротив, сыновья получили от
отца Y-хромосому, в которой нет интересующих
нас локусов, и их фенотип целиком
зависит от того, какие гены получили
они с Х-хромосомой матери. По расщеплению
у сыновей нетрудно видеть, что их мять
гетерозиготна по обоим генам
(CcDd).
Чтобы сказать, как комбинируются эти
гены в двух Х-хромосомах женщины, нужно
использовать сведения о ее родителях.
Ее отец страдал мышечной дистрофией,
но имел нормальное зрение. Следовательно
женщина получила от своего отца его
единственную Х-хромосому с генами
Cd.
А поскольку она дигетерозиготна, она
должна была получить от своей матери
Х-хромосому с генами —
cD.
Таким образом, генотипы жены и мужа
могут быть представлены формулами:
О
числе кроссоверных и некроссоверных
гамет женщины можно судить по фенотипу
ее сыновей. Из некроссоверных гамет
развились: 3 сына с дистрофией и 3 с
цветовой слепотой (всего 6). Из кроссоверных
— 1 имеющий обе патологи и 1 здоровый (2
рекомбинанта). Теперь, для определения
расстояния между генами
cud
узнаем, какой процент составляют
кроссоверные гаметы из общего числа
гамет, давших сыновей:
100
х 2/8= 25 морганид
Оценка
должна рассматриваться как приблизительная,
в связи с малым числом особей, по которым
произведено вычисление.
Разберем
задачу 107 а). В задаче даны расстояния
между генами, локализованными в одной
и той же хромосоме. Расстояние
LM
равно 5 морганид,
a MN
— 3 морганиды. Определить расстояние
между
L
и N.
Решение
этой задачи основывается на законе
линейного расположения генов в хромосоме
(закон Моргана). Если три точки (А, В и С)
расположены на прямой линии, то расстояние
АС равно либо сумме, либо разности
расстояний АВ и ВС.
АС
= АВ ± ВС
Эта
формула, являющаяся количественным
выражением закона Моргана, позволяет
дать два возможных варианта
взаиморасположения трех точек на
прямой (см. схему 43).
АВ+ВС=5+3
А В С__
АВ—ВС=5-3
А С В_________
Схема
43. Определение расстояния между тремя
генами
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
07.03.201629.8 Mб35Генетика Лобашев М Е 1967г.djvu
- #
- #
- #
Сцепленное наследование
02-Сен-2014 | Нет комментариев | Лолита Окольнова
Сцепленное наследование
Автор статьи — Саид Лутфуллин.
После открытия законов наследственности Менделя стали замечать, что не всегда эти законы срабатывают.
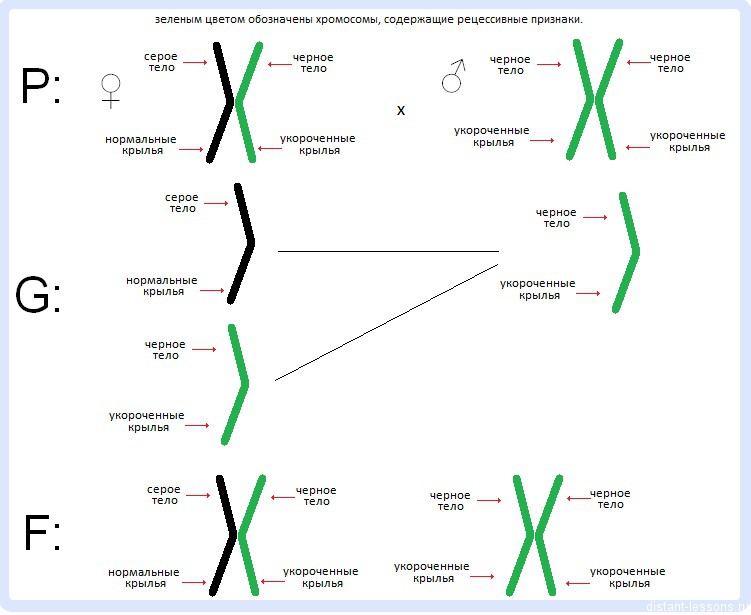
Например: скрестили дигетерозиготную самку дрозофилы с серым телом и нормальными крыльями с самцом с черным телом и укороченными крыльями.
Серое тело и нормальные крылья – доминантные признаки.
По законам Менделя схема скрещивания такая:

Но практический результат скрещивания отличается.
Как правило, в потомстве наблюдается расщепление 1:1,
фенотипы потомства: серое тело, нормальные крылья и черное тело, укороченные крылья.
Не срабатывает закон независимого наследования. Почему же так? Неужели законы Менделя действительно не работают? Конечно же, нет, законы природы, могут быть «нарушены», только если это позволяет другой закон (исключение из правила).
Давайте разберемся…
- информацию о каждом признаке несет определенный ген;
- гены находятся в хромосомах.
Естественно, что количество хромосом значительно меньше количества генов, поэтому в одной хромосоме закодировано несколько генов.
Гены, находящиеся в одной хромосоме наследуются вместе, то есть сцеплено.
А гены, находящиеся в разных хромосомах наследуются независимо,
так как при гаметогенезе хромосомы распределяются случайно, следовательно, два несцепленных гена могут попасть вместе в одну гамету гамете, а могут и нет.
Гены, находящиеся в одной хромосоме, обязательно окажутся в одной гамете.
В примере, который мы рассмотрели ранее, мы можем заметить: серое тело наследуется вместе с нормальные крыльями, а черное тело наследуется вместе с укороченными крыльями.
Гены цвета тела и длины крыльев находятся в одной хромосоме.

Самка дигетерозиготна, есть две гомологичные хромосомы:
в одной из гомологичных хромосом закодированы гены серого тела и нормальных крыльев,
в другой — гены черного тела и укороченных крыльев
Но получается всего два вида гамет — признаки цвета тела и размера крыльев «неделимы»
Отцовская особь по этим признакам дигомозиготная:
в одной гомологичной хромосоме гены черного тела и укороченных крыльев,
и в другой гомологичной хромосоме так же.
Все признаки, закодированные в одной хромосоме, образуют так называемую группу сцепления.
Признаки из одной группы сцепления наследуются вместе.
И как можно догадаться,
количество групп сцепления равно количеству хромосом в гаплоидном наборе.
Примеры задач
Задача 1:
Немного другое оформление: сцепленные признаки записываются на «палочках», например генотип самки из нашей задачи следует записать вот так:
![]()
- палочки означают гомологичные хромосомы, в которых локализованы гены
- буквы по одну сторону от палочек обозначают сцепленные друг с другом гены.
То есть запись говорит:
признаки АВ сцеплены друг с другом; признаки ab так же сцеплены друг с другом
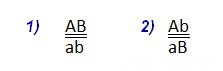
- положение генов в генотипе 1) называется цис-положением: AB \ ab (доминантные признаки на одной хромосоме, рецессивные на другой)
- положение 2) называется транс-положением: Ab \ aB.
Разберем на примере:

1) В условии задачи сразу указаны все признаки, заполним таблицу:
|
Ген |
Признак |
|
А |
|
|
а |
|
|
B |
Усики есть |
|
b |
Усиков нет |
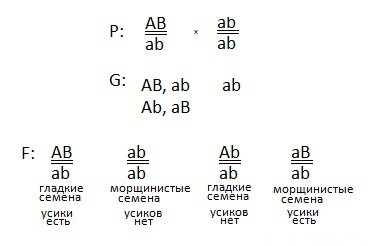
2) Первое растение дигетерозиготно, сказано, что доминантные признаки локализованы в одной хромосоме, то есть сцеплены. Причем доминантные признаки находятся на одной гомологичной хромосоме, следовательно на другой гомологичной хромосоме располагаются рецессивные признаки (цис-положение). Генотип первого растения: AB \ ab.
Получаем всего два вида гамет (так как признаки сцеплены):
AB и ab.
3) Так как у второго растения проявились рецессивные признаки, делаем вывод, что оно дигомозиготно. И его генотип: ab\ab. Образуется только один сорт гамет: ab.
4) Наконец, составим схему скрещивания:
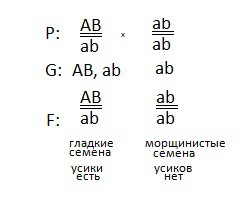
И ответим на последний вопрос задачи — про закон:
проявляется закон сцепленного наследования, он гласит: гены, локализованные в одной хромосоме образуют группу сцепления и наследуются вместе.
Но случается, что даже гены из одной группы сцепления (локализованные в одной хромосоме) наследуются раздельно, то есть «расцепляются».
Для примера, возьмем скрещивание из предыдущей задачи.
При таком же скрещивании может получиться и 4 фенотипические группы (вместо положенных 2) в потомстве, как и при независимом наследовании. Это объясняется возможностью кроссинговера между гомологичными хромосомами (тем, кто не понимает о чем речь, советую прочитать статью кроссинговер).
Допустим если у особи признаки AB сцеплены, то при образовании гамет, если произойдет кроссинговер, есть вероятность, что участок хромосомы, в котором закодирован один из генов «перескочит» на другую гомологичную хромосому, и сцепление нарушится. На примере нашей задаче, в случае кроссинговера скрещивание будет следующим:
У дигетерозиготного растения образуется еще два сорта гамет, за счет кроссинговера. Гаметы, при образовании которых, произошел кроссинговер (в данной задаче это Ab и aB) называются кроссоверными. Статистически процент кроссоверных гамет меньше некроссоверных.
Чем дальше друг от друга находятся гены в хромосоме, тем больше вероятность того, что сцепленные гены, будут «разлучены» при рекомбинации, происходящей во время кроссинговера.
И соответственно, чем ближе друг к другу расположены гены в хромосоме, тем вероятность их разъединения.
Эта зависимость вероятности разделения генов кроссинговером и расстояния между генами оказалась настолько «удобной», что расстояние между генами измеряют в процентах вероятности их разъединения при кроссинговере. По формуле:
![]()
Где:
- x – вероятность разъединения генов в процентах,
- а – количество особей, образовавшихся из кроссоверных гамет, n – количество всех особей.
- И 1% вероятности разъединения генов приняли за единицу расстояния между этими генами.
Единица эта называется морганида. Назвали единицу в честь известного генетика Томаса Моргана, который изучал это явление
1 морганида = 1% вероятности, что сцепленные гены, в результате кроссинговера, окажутся на разных гомологичных хромосомах
Задача 2:

1) Составим таблицу признаков
|
Ген |
Признак |
|
А |
|
|
а |
|
|
B |
Гладкий эндосперм |
|
b |
Шероховатый эндосперм |
2) Раз в условии сказано, что провели анализирующее скрещивание, значит второе растение дигомозиготно по рецессивным признакам, его генотип: ab \ ab.
3) В потомстве получили 4 фенотипических группы. Так как спризнаки сцеплены, то очевидно имел место кроссинговер. Так же появление четырех фенотипических групп при анализирующем скрещивании указывает на дигетерозиготность первого растения. Значит, его генотип: либо AB \ ab, либо Ab \ aB.
Чтобы определить в каком положении гены — цис или транс, надо посмотреть на соотношение в потомстве. Процент кроссоверных гамет меньше некроссоверных, поэтому особей, получившихся из некроссоверных гамет больше.
Эти особи: 208 высоких растений с гладким эндоспермом, 195 – низких с шероховатым эндоспермом
У них доминантный признак унаследовался с доминантным, а рецессивный с рецессивным. Следовательно гены у родительской дигетерозиготной особи в цис-положении: AB \ ab.
4) Схема скрещивания:
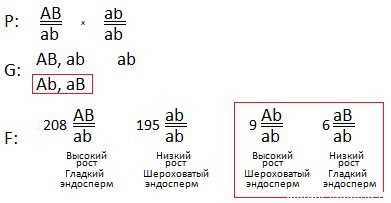
Красным отмечены кроссоверные гаметы и особи, полученные из них. Этих особей меньше, так как кроссоверных гамет образуется меньше. Если бы у родительской дигетерозиготной особи гены были бы в транс положении, в потомстве наоборот образовалось бы больше особей с признаками: высоких шероховатых, и низких гладких.
5) Определим расстояние между генами.
Для этого вычислим вероятность того, что признаки окажутся разделенными в результате кроссинговера.
По формуле:
![]()
x= 9 + 6208+ 195+ 9 + 6 ×100%= 15418 ×100%=3,59%
То есть расстояние между генами = 3,59 морганид.
Обсуждение: “Сцепленное наследование”
(Правила комментирования)
