From Wikipedia, the free encyclopedia
Water potential is the potential energy of water per unit volume relative to pure water in reference conditions. Water potential quantifies the tendency of water to move from one area to another due to osmosis, gravity, mechanical pressure and matrix effects such as capillary action (which is caused by surface tension). The concept of water potential has proved useful in understanding and computing water movement within plants, animals, and soil. Water potential is typically expressed in potential energy per unit volume and very often is represented by the Greek letter ψ.
Water potential integrates a variety of different potential drivers of water movement, which may operate in the same or different directions. Within complex biological systems, many potential factors may be operating simultaneously. For example, the addition of solutes lowers the potential (negative vector), while an increase in pressure increases the potential (positive vector). If the flow is not restricted, water will move from an area of higher water potential to an area that is lower potential. A common example is water with dissolved salts, such as seawater or the fluid in a living cell. These solutions have negative water potential, relative to the pure water reference. With no restriction on flow, water will move from the locus of greater potential (pure water) to the locus of lesser (the solution); flow proceeds until the difference in potential is equalized or balanced by another water potential factor, such as pressure or elevation.
Components of water potential[edit]
Many different factors may affect the total water potential, and the sum of these potentials determines the overall water potential and the direction of water flow:
 [1]
[1]

where:
All of these factors are quantified as potential energies per unit volume, and different subsets of these terms may be used for particular applications (e.g., plants or soils). Different conditions are also defined as reference depending on the application: for example, in soils, the reference condition is typically defined as pure water at the soil surface.
Pressure potential[edit]
Pressure potential is based on mechanical pressure and is an important component of the total water potential within plant cells. Pressure potential increases as water enter a cell. As water passes through the cell wall and cell membrane, it increases the total amount of water present inside the cell, which exerts an outward pressure that is opposed by the structural rigidity of the cell wall. By creating this pressure, the plant can maintain turgor, which allows the plant to keep its rigidity. Without turgor, plants will lose structure and wilt.
The pressure potential in a plant cell is usually positive. In plasmolysed cells, pressure potential is almost zero. Negative pressure potentials occur when water is pulled through an open system such as a plant xylem vessel. Withstanding negative pressure potentials (frequently called tension) is an important adaptation of the xylem. This tension can be measured empirically using the Pressure bomb.
Osmotic potential (solute potential)[edit]
Pure water is usually defined as having an osmotic potential ( ) of zero, and in this case, solute potential can never be positive. The relationship of solute concentration (in molarity) to solute potential is given by the van ‘t Hoff equation:
) of zero, and in this case, solute potential can never be positive. The relationship of solute concentration (in molarity) to solute potential is given by the van ‘t Hoff equation:

where  is the concentration in molarity of the solute,
is the concentration in molarity of the solute,  is the van ‘t Hoff factor, the ratio of amount of particles in solution to amount of formula units dissolved,
is the van ‘t Hoff factor, the ratio of amount of particles in solution to amount of formula units dissolved,  is the ideal gas constant, and
is the ideal gas constant, and  is the absolute
is the absolute
temperature.

The water diffuses across the osmotic membrane to where the water potential is lower
For example, when a solute is dissolved in water, water molecules are less likely to diffuse away via osmosis than when there is no solute. A solution will have a lower and hence more negative water potential than that of pure water. Furthermore, the more solute molecules present, the more negative the solute potential is.
Osmotic potential has important implications for many living organisms. If a living cell is surrounded by a more concentrated solution, the cell will tend to lose water to the more negative water potential ( ) of the surrounding environment. This can be the case for marine organisms living in sea water and halophytic plants growing in saline environments. In the case of a plant cell, the flow of water out of the cell may eventually cause the plasma membrane to pull away from the cell wall, leading to plasmolysis. Most plants, however, have the ability to increase solute inside the cell to drive the flow of water into the cell and maintain turgor.
) of the surrounding environment. This can be the case for marine organisms living in sea water and halophytic plants growing in saline environments. In the case of a plant cell, the flow of water out of the cell may eventually cause the plasma membrane to pull away from the cell wall, leading to plasmolysis. Most plants, however, have the ability to increase solute inside the cell to drive the flow of water into the cell and maintain turgor.
This effect can be used to power an osmotic power plant.[2]
A soil solution also experiences osmotic potential. The osmotic potential is made possible due to the presence of both inorganic and organic solutes in the soil solution. As water molecules increasingly clump around solute ions or molecules, the freedom of movement, and thus the potential energy, of the water is lowered. As the concentration of solutes is increased, the osmotic potential of the soil solution is reduced. Since water has a tendency to move toward lower energy levels, water will want to travel toward the zone of higher solute concentrations. Although, liquid water will only move in response to such differences in osmotic potential if a semipermeable membrane exists between the zones of high and low osmotic potential. A semipermeable membrane is necessary because it allows water through its membrane while preventing solutes from moving through its membrane. If no membrane is present, movement of the solute, rather than of the water, largely equalizes concentrations.
Since regions of soil are usually not divided by a semipermeable membrane, the osmotic potential typically has a negligible influence on the mass movement of water in soils. On the other hand, osmotic potential has an extreme influence on the rate of water uptake by plants. If soils are high in soluble salts, the osmotic potential is likely to be lower in the soil solution than in the plant root cells. In such cases, the soil solution would severely restrict the rate of water uptake by plants. In salty soils, the osmotic potential of soil water may be so low that the cells in young seedlings start to collapse (plasmolyze).
Matrix potential (Matric potential)[edit]
When water is in contact with solid particles (e.g., clay or sand particles within soil), adhesive intermolecular forces between the water and the solid can be large and important. The forces between the water molecules and the solid particles in combination with attraction among water molecules promote surface tension and the formation of menisci within the solid matrix. Force is then required to break these menisci. The magnitude of matrix potential depends on the distances between solid particles—the width of the menisci (also capillary action and differing Pa at ends of the capillary)—and the chemical composition of the solid matrix (meniscus, macroscopic motion due to ionic attraction).
In many cases, the absolute value of matrix potential can be relatively large in comparison to the other components of water potential discussed above. Matrix potential markedly reduces the energy state of water near particle surfaces. Although water movement due to matrix potential may be slow, it is still extremely important in supplying water to plant roots and in engineering applications. The matrix potential is always negative because the water attracted by the soil matrix has an energy state lower than that of pure water. Matrix potential only occurs in unsaturated soil above the water table. If the matrix potential approaches a value of zero, nearly all soil pores are completely filled with water, i.e. fully saturated and at maximum retentive capacity. The matrix potential can vary considerably among soils. In the case that water drains into less-moist soil zones of similar porosity, the matrix potential is generally in the range of −10 to −30 kPa.
Empirical examples[edit]
Soil-plant-air continuum[edit]
At a potential of 0 kPa, soil is in a state of saturation. At saturation, all soil pores are filled with water, and water typically drains from large pores by gravity. At a potential of −33 kPa, or −1/3 bar, (−10 kPa for sand), soil is at field capacity. Typically, at field capacity, air is in the macropores, and water in the micropores. Field capacity is viewed as the optimal condition for plant growth and microbial activity. At a potential of −1500 kPa, the soil is at its permanent wilting point, at which plant roots cannot extract the water through osmotic diffusion. Soil waterways still evaporate at more negative potentials down to a hygroscopic level, at which soil water is held by solid particles in a thin film by molecular adhesion forces.
In contrast, atmospheric water potentials are much more negative—a typical value for dry air is −100 MPa, though this value depends on the temperature and the humidity. Root water potential must be more negative than the soil, and the stem water potential must be an intermediate lower value than the roots but higher than the leaf water potential, to create a passive flow of water from the soil to the roots, up the stem, to the leaves and then into the atmosphere.[3][4][5]
Measurement techniques[edit]
A tensiometer, electrical resistance gypsum block, neutron probes, or time-domain reflectometry (TDR) can be used to determine soil water potential energy. Tensiometers are limited to 0 to −85 kPa, electrical resistance blocks are limited to −90 to −1500 kPa, neutron probes are limited to 0 to −1500 kPa, and a TDR is limited to 0 to −10,000 kPa. A scale can be used to estimate water weight (percentage composition) if special equipment is not on hand.
See also[edit]
- Water retention curve
- Pore water pressure
Notes[edit]
- ^ Taiz; Zeiger (2002). Plant Physiology (Fourth ed.). Sinauer Associates.
- ^ “Statkraft to build world’s first osmotic power plant”. Archived from the original on 2009-02-27. Retrieved 2014-01-29.
- ^ Beerling, D. J. (2015). “Gas valves, forests and global change: a commentary on Jarvis (1976) ‘The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field’“. Philosophical Transactions of the Royal Society B: Biological Sciences. 370 (1666): 20140311. doi:10.1098/rstb.2014.0311. ISSN 0962-8436. PMC 4360119. PMID 25750234.
- ^ Jarvis, P. G. (1976). “The Interpretation of the Variations in Leaf Water Potential and Stomatal Conductance Found in Canopies in the Field”. Philosophical Transactions of the Royal Society B: Biological Sciences. 273 (927): 593–610. Bibcode:1976RSPTB.273..593J. doi:10.1098/rstb.1976.0035. ISSN 0962-8436.
- ^ Jones, Hamlyn G. (2013-12-12). Plants and Microclimate: A Quantitative Approach to Environmental Plant Physiology. Cambridge University Press. p. 93. ISBN 9781107511637.
External links[edit]
- http://lawr.ucdavis.edu/classes/ssc107/SSC107Syllabus/chapter2-00.pdf
Водный потенциал клетки (Схема)
Из неорганических веществ, которые входят в состав клетки, самым важным является вода. Количество воды в общей массы организма 60% до 95%.
Водный потенциал системы схема
Водный потенциал – это мера свободной кинетической энергии воды в системе, или стремления воды выйти за пределы системы. Он измеряется в единицах давления (кПа) и обозначается символом Ψ («пси»).
Водный потенциал чистой воды приравнивается к нулю – это точка отсчета, как и в системе окислительно-восстановительного потенциала, используемой в химии.
т.е. для чистой воды Ψ = 0
Присутствие в растворе молекул растворенного вещества ослабляет тенденцию молекул воды к перемещению из раствора.
Следовательно Ψраствора < 0
(молекулы растворенного вещества в растворе «мешают» движению молекул воды, следовательно, кинетическая энергия молекул воды снижена и Ψ становится отрицательным).
Вода движется по градиенту водного потенциала, т.е. из области с менее отрицательным потенциалом (например, -500 кПа) в область с более отрицательным (например, -1000 кПа).

Преимущества системы водного потенциала:
1) движение воды рассматривается скорее с точки зрения «системы», чем с точки зрения окружающей среды;
2) можно проводить сравнения между разными системами, например, между атмосферой, воздухом, содержащимся в листе, и клетками мезофила листа.
Необходимо помнить, что: Осмос – это движение воды через полупроницаемую мембрану по градиенту водного потенциала.
Водный потенциал клетки
Присутствие в клеточном соке растворенных веществ снижает Ψр (делает его более отрицательным). Поскольку для чистой воды Ψ = 0, если поместить в нее растительную клетку то вода, двигаясь по градиенту водного потенциала, будет поступать в клетку.
Целлюлозная клеточная стенка свободно проницаема для воды и растворенных веществ.
Плазматическая мембрана свободно проницаема для воды, но частично проницаема для растворенных веществ.
Поступление воды в клетку приведет к набуханию протопласта (содержимого клетки, окруженного плазматической мембраной). В результате усилится давление на клеточную стенку (тургорное давление). Расширению протопласта противостоит сила, равная тургорному давлению, но направленная противоположно ему. Она называется потенциалом давления (Ψд).
Потенциал давления стремится противостоять поступлению воды или выталкивать воду из клетки.
Ψклетки = Ψр + Ψд
где:
Ψклетки – это водный потенциал клетки, т.е. стремление воды покинуть клетку.
Ψр – это влияние концентрации растворенного вещества на осмотический потенциал – это всегда отрицательная величина.
Ψд – это влияние потенциала давления и тургорного давления на водный потенциал представляет стремление воды к выходу из клетки. Эта величина либо равна нулю, либо имеет положительное значение.
Пример расчета водного потенциала клетки
Движение воды между клетками А и Б:

Водный потенциал клетки А: Ψ = -1300 + 500 = -800 кПа
Водный потенциал клетки Б: Ψ = -1900 + 700 = -1200 кПа
Поскольку число – 8 0 0 больше, чем -1 2 0 0 , вода будет двигаться по градиенту водного потенциала из А в Б.
_______________
Источник информации:
Биология человека в диаграммах / В.Р. Пикеринг — 2003.
Водный потенциал
Водный
потенциал —
один из фундаментальных показателей
термодинамического состояния воды. Ее
молекулы обладают кинетической энергией,
поскольку и в жидкости, и в газе (водяном
паре) быстро и беспорядочно движутся.
Чем выше концентрация этих молекул в
системе, тем больше их общая кинетическая
энергия и выше водный потенциал. Таким
образом, у чистой воды он максимален.
Если две системы, содержащие воду
(например, почва и атмосфера или клетка
и окружающий ее раствор), вступают в
контакт, то беспорядочное движение ее
молекул приведет к их результирующему
движению туда, где их меньше, — из области
с более высоким водным потенциалом (с
более высокой энергией) туда, где он
ниже (ниже энергия), пока их концентрации
в обеих системах не уравновесятся.
В
биологии водный потенциал обычно
измеряют единицами давления (например,
Паскалями)1. У чистой воды он максимален,
но условно принят за нулевой.
Запомните
следующие главные моменты:
1)
водный потенциал чистой воды максимален
и, по определению, равен нулю (в
«стандартных» условиях, т. е. при
нормальном атмосферном давлении и 25
°С);
2)
вода всегда движется по градиенту у:
оттуда, где он выше, туда, где он ниже;
3)
у всех растворов водный потенциал ниже,
чем у чистой воды, т. е. в стандартных
условиях выражается отрицательной
величиной;
4)
осмос можно определить как движение
молекул воды через полупроницаемую
мембрану по градиенту водного потенциала.
Преимущество использования понятия «водный потенциал»
Водный
потенциал отражает
тенденцию молекул воды двигаться в
определенном направлении. Чем он выше
(чем менее отрицателен), тем больше у
молекул шансов покинуть данную систему.
Если две системы (не обязательно
разделенные мембраной) контактируют
друг с другом, то вода будет переходить
из системы с более высоким водным
потенциалом в систему с более низким
водным потенциалом.
Таким
образом, понятие «водный
потенциал»
позволяет определить направление
движения воды между любыми двумя
системами — не только из одной клетки
растения в другую, но, например, из почвы
в корень, из листа в атмосферу и т. п.
Можно сказать, что вода движется через
растение по градиенту водного потенциала
из почвы в воздух. Чем круче этот градиент,
тем быстрее поток воды.
17
Проницаемость и свойства клеточных мембран
Разные
вещества проходят через мембрану с
разной скоростью, поэтому мы говорим,
что мембраны избирательно проницаемы.
При этом скорость прохождения веществ
меняется в зависимости от физиологического
состояния клетки или органеллы.
Благодаря
избирательной проницаемости они
регулируют транспорт веществ между
наружной средой и клеткой, между
органеллами цитоплазмой и т. д.
Регулируя
поступление веществ в клетку и их
выведение, мембраны тем самым регулируют
скорость и направление биохимических
реакций, которые составляют основу
обмена веществ организма. Сама
избирательная проницаемость мембран
зависит от обмена веществ в клетке.
Мембраны
регулируют обмен веществ и другим
способом – изменяя активность ферментов.
Некоторые ферменты активны только
тогда, когда они прикреплены к мембране,
другие, наоборот, в этом состоянии не
проявляют активности и начинают
действовать только после того, как
мембрана выпустит их на «свободу».
Изменение проницаемости мембраны может
способствовать контакту фермента с
субстратом, после чего начинается
химическая реакция, которая сначала
была невозможна.
Мембранные
ферменты работают хорошо только тогда,
когда они находятся в контакте с липидами.
В присутствии липидов может меняться
форма молекул мембранных белков –
ферментов, таким образом, что их активные
центры становятся доступным для
субстрата. Кроме того, локализация
фермента на мембране определяет место
данной реакции в клетке.
Другой
важной стороной ферментативной
деятельности мембран является координация
химических реакций, проходящих в клетках.
Когда несколько ферментов катализируют
цепь реакций, в которой продукт первой
реакции служит субстратом для другой
и т. д., то эти ферменты располагаются
на мембране в определенной последовательности,
образуя мультиферментную систему. Таких
систем в мембране много, например цепь
дыхательных ферментов. В этом случае
ферменты располагаются в строгой
последовательности с минимальным
расстоянием между ними.
Компартментализация клетки –
необходимое условие для жизнедеятельности
и одна из основных функций мембран.
Во-первых, мембраны увеличивают внутреннюю
поверхность клетки, на которой локализованы
ферменты и проходят химические реакции.
Во-вторых, разные компартменты отличаются
по химическому составу. Далее, поскольку
компартменты имеют различный химический
состав в них проходят разные биохимические
реакции, то с помощью мембран осуществляется
физическое разделение метаболических
процессов, часто противоположного
направления. Например, синтез белков
идет в рибосомах, а распад – в
лизосомах. Каждый из этих процессов
регулируется независимо один от другого.
Приведем еще пример: синтез жирных
кислот и их окисление. Первый процесс
происходит в цитоплазме, второй – в
митохондриях.
Однако
метаболические системы не полностью
изолированы одна от другой. В мембранах,
разделяющих клетку на компартменты,
имеются специализированные механизмы,
которые транспортируют из одного в
другой субстраты, продукты реакции, а
также кофакторы и соединения, имеющие
регуляторное действие. Таким образом,
скорость отдельных метаболических
процессов, которые происходят внутри
компартментов, частично регулируются
транспортными системами мембран.
Регуляция
скорости метаболических процессов
может происходить благодаря перемещению
регулируемых веществ с одного компартмента
в другой.
В
разных компартментах имеются разные
концентрации органических веществ,
ионов, разный химический состав. Например,
в вакуолях всегда находится запас
аминокислот, органических кислот,
сахаров, ионов. Это приводит к химической
геторогенности в клетке. Неодинаковая
концентрация ионов по обеим сторонам
мембраны приводит к возникновению
разности электрических потенциалов.
Так плазмалемма несет отрицательный
заряд, а тонопласт – положительный.
Разные концентрации и химический состав
обуславливают разную вязкость в разных
частях цитоплазмы.
Обладая
избирательной проницаемостью, пропуская
в клетку необходимые вещества, мембраны
выполняют еще одну функцию – регулируют
гомеостаз. Гомеостазомназывают
свойство клетки (органеллы, органа,
организма, экосистемы) поддерживать
постоянство своей внутренней среды.
Почему
внутренняя среда клетки должна оставаться
постоянной? Мембранные белки и
белки-ферменты относятся к глобулярным.
Глобулярная нативная структура белковых
молекул зависит от слабых связей, легко
разрушаемым даже при малом изменении
внутренней среды клетки. Таким образом,
клетка должна поддерживать гомеостаз,
чтобы не изменялась нативная структура
белков. Если измениться третичная или
четвертичная структура белка, то и
фермент потеряет или изменит свою
активность и нарушится строгое
соответствие структуры фермента и
субстрата, для того чтобы пошла реакция.
От
структуры белковой молекулы зависит
ее размещение в мембране, и, таким
образом, ее свойства и функции. Изменение
конформации белковых молекул может
менять количество гидрофобных и
гидрофильных радикалов на ее поверхности.
Это приводит к изменению расположения
белковых глобул в мембране. Последнее
окажет влияние на ее избирательную
способность и другие свойства, что, в
свою очередь, вызовет нарушение
геторогенности, исчезновению ферментов
и может привести к гибели клетки.
Мембраны
принимают участие в адаптации клетки
к меняющимся условиям окружающей среде,
о чем поговорим ниже.
Большая
часть мембран, кроме общих функций,
таких как регулирование обмена веществ,
компартментизация, выполняют и
специальные. Например, мембраны
митохондрий и хлоропластов принимают
непосредственное участие в синтезе
АТФ. Жизнь – это беспрерывная работа,
для выполнения которой все время
необходимо расходовать энергию.
Таким
образом, синтез АТФ необходим постоянно,
он связан со строго определенной
структурой мембран органелл (хлоропласты,
митохондрии). Нарушение этой структуры
приводит к снижению синтеза АТФ, а это
значит – к смерти.
Лабильная
структура мембран позволяет им выполнять
разные функции: барьерные, транспортные
осмотические, электрические, структурные,
энергетические, биосинтетические,
секреторные, рецепторно- регуляторные
и некоторые другие.
В
последнее время накапливается все
больше данных, свидетельствующих о том,
что некоторые мембраны образуются путем
физического переноса мембранного
материала от одних клеточных компонентов
к другим. Есть данные, позволяющие
считать ЭС источником тех строительных
блоков, которые в конечном итоге
включаются в плазмалемму. Возможно, это
происходит в результате отшнуровывания
пузырьков от цистерн Гольджи. По всей
вероятности, в аппараты Гольджи
совершается перестройка мембран двух
типов: мембран, характерных для ЭС, в
мембраны, свойственные плазмалемме.
В
заключение укажем на основные свойства
мембран:
1.
Мембраны являются сложными структурами.
Они состоят из структурных белков и
липидов, но могут также включать
высокоспецифические молекулы ферментов,
пигментов и кофакторов.
2.
Благодаря химической вариабильности
составляющих мембраны молекул белков
и липидов и в зависимости от их функций,
различные мембраны могут иметь разную
структуру.
3.
Структура мембран обеспечивает высокую
степень упорядоченности которой
специфические молекулы могут образовывать
комплексные функциональные единицы.
4.
Ферментные реакции и другие процессы
в мембранах могут приводить к
пространственно направленным, или
векторным, реакциям; мембраны асимметричны
Плазмолиз
(от греч. plásma — вылепленное, оформленное
и lýsis — разложение, распад) , отделение
протопласта от оболочки при погружении
клетки в гипертонический раствор.
Плазмолиз
характерен главным образом для
растительных клеток, имеющих прочную
целлюлозную оболочку. Животные клетки
при перенесении в гипертонический
раствор сжимаются. В зависимости от
вязкости протоплазмы, от разницы между
осмотическим давлением клетки и внешнего
раствора, а следовательно от скорости
и степени потери воды протоплазмой,
различают плазмолиз выпуклый, вогнутый,
судорожный и колпачковый. Иногда
плазмолизированные клетки остаются
живыми; при погружении таких клеток в
воду или гипотонический раствор
происходит деплазмолиз.
Для
сравнительной оценки плазмолиза в
тканях существует два метода:
–
Метод пограничного плазмолиза
–
Плазмометрический метод.
Первый
метод, разработанный Хуго Де Фризом
(1884), заключается в погружении тканей в
растворы с различной концентрацией
KNO3, сахарозы или другие осмотически
активного вещества и установлении той
концентрации, при которой плазмолизируется
50 % клеток. При плазмометрическом методе
после плазмолиза измеряют относительный
объём клетки и протопласта и по
концентрации раствора вычисляют
осмотическое давление клетки (по
соответствующим формулам) .
Деплазмолиз
(от де… и плазмолиз) — возвращение
протопласта клеток растений из состояния
плазмолиза в исходное состояние,
характеризующееся нормальным тургором.
Деплазмолиз
происходит при перенесении плазмолизированных
клеток (то есть клеток, подвергшихся
плазмолизу) в воду или гипотонические
растворы.
Тургор
(позднелат. turgor — вздутие, наполнение,
от лат. turgere — быть набухшим, наполненным)
, напряжённое состояние клеточной
оболочки, зависящее от осмотического
давления внутриклеточной жидкости (Р
внутреннее) , осмотическое давления
внешнего раствора (Р внешнее) и упругости
клеточной оболочки (УО) . Обычно УО клеток
животных (исключая некоторых
кишечнополостных) невелика, они лишены
высокого Т. и сохраняют целостность
только в изотонических растворах или
мало отличающихся от изотонических
(разница между Р внутренним и Р внешним
меньше 0,5—1,0 ам) . У живых растительных
клеток Р внутреннее всегда больше Р
внешнего, однако разрыва клеточной
оболочки у них не происходит из-за
наличия целлюлозной клеточной стенки.
Разница между Р внутренним и Р внешним
у растений (например, у растений галофитов,
грибов) достигает 50—100 ам, но даже при
этом запас прочности растительной
клетки составляет 60—70%. У большинства
растений относительное удлинение
клеточной оболочки вследствие Т. не
превышает 5— 10%, а тургорное давление
лежит в пределах 5—10 ам. Благодаря Т.
ткани растений обладают упругостью и
конструктивной прочностью. Все процессы
автолиза, увядания и старения сопровождаются
падением Т.
18
Вода́ (оксид водорода) — бинарное неорганическое
соединение, химическая
формула Н2O.
Молекула воды состоит из двух
атомовводорода и
одного — кислорода,
которые соединены между собой ковалентной
связью.
При нормальных
условиях представляет
собой прозрачную жидкость,
не имеет цвета (в
малом объёме), запаха и вкуса.
В твёрдом состоянии называется льдом (кристаллы
льда могут образовывать снег или иней),
а в газообразном —
водяным паром.
Вода также может существовать в
виде жидких
кристаллов (нагидрофильных поверхностях)[3][4].
Около 71 % поверхности Земли покрыто
водой (океаны, моря, озёра, реки,
льды) — 361,13 млн км2[5][6].
На Земле примерно 96,5 % воды приходится
на океаны, 1,7 % мировых запасов
составляют грунтовые воды, ещё 1,7 %
на ледники и ледяные шапки Антарктиды и Гренландии,
небольшая часть в реках, озёрах и болотах,
и 0,001 % в облаках (образуются из
взвешенных в воздухе частиц льда и
жидкой воды)[7][8].
Бо́льшая часть земной воды — солёная,
и она непригодна для сельского
хозяйства и
питья. Доляпресной составляет
около 2,5 %, причём 98,8 % этой воды
находится в ледниках и грунтовых
водах.
Менее 0,3 % всей пресной воды содержится
в реках, озёрах и атмосфере,
и ещё меньшее количество (0,003 %)
находится в живых организмах[7].
Является
хорошим сильнополярным растворителем.
В природных условиях всегда содержит
растворённые вещества (соли, газы).
Вода
имеет ключевое значение в создании и
поддержании жизни на
Земле, в химическом строении живых
организмов,
в формированииклимата и погоды.
Является важнейшим веществом для всех
живых существ на планете Земля[9].
Первая
особенность: вода – единственное
вещество на Земле (кроме ртути),
для
которого зависимость удельной теплоемкости
от температуры имеет
минимум.Из-за
того, что удельная теплоемкость воды
имеет
минимум около 37°С, нормальная
температура человеческого тела,
состоящего
на две трети из воды, находится в диапазоне
температур
36°-38°С(внутренние органы
имеют более высокую температуру,
чем
наружные).
Вторая
особенность: теплоемкость воды
аномально
высока. Чтобы нагреть
определенное ее количество на один
градус,
необходимо затратить больше
энергии, чем при нагреве других жидкостей,
–
по крайней мере вдвое по отношению
к простым веществам. Из этого
вытекает
уникальная способность воды сохранять
тепло. Подавляющее
большинство
других веществ таким свойством не
обладают. Эта
исключительная
особенность воды способствует тому,
что у человека
нормальная температура
тела поддерживается на одном уровне и
жарким
днем, и прохладной
ночью.
Таким образом, вода
играет
главенствующую роль в
процессах регулирования теплообмена
человека и
позволяет ему поддерживать
комфортное состояние при
минимуме
энергетических затрат.
При нормальной температуре тела человек
находится
в наиболее выгодном
энергетическом состоянии.
Температура
других
теплокровных млекопитающих (32-39°С) также
хорошо соотносится с
температурой
минимума удельной теплоемкости
воды.
Третья
особенность: вода
обладает высокой удельной теплотой
плавления, то есть
воду очень трудно
заморозить, а лед – растопить. Благодаря
этому климат
на Земле в целом
достаточно стабилен и мягок.
Все
три особенности тепловых свойств воды
позволяют человеку оптимальным
образом
существовать в условия благоприятной
среды.
выполняет
транспортную функцию по «доставке»
питательных веществ тканям и
органам
при корневом и листовом питании, обмеенных
процессах и синтезе,
–
термолегулирующую, препятствующую
перегреву тканей и денатурации
(разрушению)
белков, в т. ч. ферментов и гормонов,
–
является основной составляющей частью
растительных организмов ( на 80-90%
растения
состоят из воды) , создающая тургор-
упругость тканей,
–
как источник элемента питания- водорода
( Н) , необходимого в процессах
фотосинтеза
первичных сахаров
19
Растительные
клетки только на самой ранней стадии
развития бывают сплошь заполнены
протоплазмой. Очень скоро в протоплазме
начинают появляться полости, вакуоли
– резервуары с клеточным соком.
Образование вакуолей обусловлено
наличием в протоплазме веществ, сильно
притягивающих воду. По мере роста и
старения клетки отдельные вакуоли
сливаются в одну сплошную полость, а
протоплазма низводится до тонкого
слоя, выстилающего клеточные стенки.
Только тяжи и нити протоплазмы пересекают
разросшуюся во всю клетку вакуолю.
Клеточный
сок, находящийся в вакуолях, имеет
сложный химический состав. В нем
содержатся в растворенном виде минеральные
соли, органические кислоты (щавелевая,
яблочная, лимонная, виннокаменная)
и их соли, сахара, азотистые вещества,
алкалоиды, глюкозиды, дубильные вещества
и др.
В
клеточном соке нередко встречаются
красящие вещества – пигменты
(антоциан, реже антохлор). Окраска
антоциана меняется в зависимости от
реакции среды. При
кислой она красная или фиолетовая, при
щелочной – синяя.
Антоцианом
окрашены корни свеклы, листья красной
капусты, фиолетовые, красные и синие
лепестки цветков. Второй растворимый
пигмент антохлор тоже иногда встречается
в лепестках и окрашивает их в желтый
цвет.
От
состава клеточного сока зависит
полезность многих культурных растений.
Сахаристость сахарной свеклы, сладкий
вкус арбуза и фруктов определяются
клеточным соком. Живая клетка растений
представляет собой осмотическую систему,
где различные вещества направляются
через мембраны от большей концентрации
к меньшей до уравнивания их.
Когда
клетка находится в воде или в очень
слабом растворе солей (как почвенный
раствор), вода поступает в клеточный
сок, вследствие чего вакуоля увеличивается
в объеме, растягивает протоплазму и
плотно прижимает ее к оболочке. Несколько
растягивается и оболочка и находится,
как говорят, в состоянии тургора
(напряжения). При большом содержании
в клетках сахара (плоды вишни, черешни,
винограда) и обильном увлажнении (частые
дожди) тургор может быть настолько
большим, что клетки лопаются.
Обратное
явление наблюдается при плазмолизе.
Если живую растительную клетку
поместить в гипертонический раствор
сахара или соли (более крепкий, чем
клеточный сок), то вода будет выходить
из клетки наружу, так как осмотическая
(притягивающая) сила такого раствора
больше осмотической силы клеточного
сока.
Особенно
велико осмотическое давление у растений,
произрастающих в пустынях и на
солончаках. Во многих случаях оно
достигает 50 и даже 100 атм. атм). По
количественным показателям, основанным
на концентрации, осмотическое давление
у некоторых растений во много раз
превышает давление пара в самых мощных
локомотивах. В действительности клеткам
приходится испытывать лишь разницу
осмотических давлений клеточного сока
и почвенных растворов, концентрация
которых в почвах пустынь и солончаках
большая.
20
Процесс
поступления веществ в клетку называется
эндоцитозом. Различают пиноцитоз и
фагоцитоз.
Фагоцитоз
(греч. фаго – пожирать) – поглощение
клеткой твердых органических веществ
. Оказавшись около клетки, твердая
частица окружается выростами мембраны,
или под ней образуется впячивание
мембраны. В результате частица оказывается
заключенной в мембранный пузырек внутри
клетки. Такой пузырек называют фагосомой.
Термин «фагоцитоз» был предложен И. И.
Мечниковым в 1882 г. Фагоцитоз свойствен
простейшим, кишечнополостным, лейкоцитам,
а также клеткам капилляров костного
мозга, селезенки, печени, надпочечников.
Второй
способ поступления веществ в клетку
называют пиноцитозом (греч. пино – пью)
– это процесс поглощения клеткой мелких
капель жидкости с растворенными в ней
высокомолекулярными веществами.
Осуществляется путем захвата этих
капель выростами цитоплазмы. Захваченные
капли погружаются в цитоплазму и там
усваиваются. Явление пиноцитоза
свойственно животным клеткам и
одноклеточным простейшим.
Еще
один способ поступления веществ в клетку
– осмос – прохождение воды через
избирательно проницаемую мембрану
клетки. Вода переходит из менее
концентрированного раствора в более
концентрированный. Вещества могут также
проходить через мембрану путем диффузии
– так транспортируются вещества,
способные растворяться в липидах
(простые и сложные эфиры, жирные кислоты
и т. д.) . Путем диффузии по градиенту
концентрации по специальным каналам
мембраны идут некоторые ионы (например,
ион калия выходит из клетки) .
Кроме
того, транспорт веществ через мембрану
осуществляет натрий-калиевый насос: он
перемещает ионы натрия из клетки и ионы
калия в клетку против градиента
концентраций с затратой энергии АТФ.
Фагоцитоз,
пиноцитоз и натрий-калиевый насос –
это примеры активного транспорта, а
осмос и диффузия – пассивного транспорта
21
ВОДНЫЙ
БАЛАНС РАСТЕНИЙ
— соотношение
между количеством воды, которое растения
получают, и количеством воды, которое
они за тот же период времени расходуют.
Водный
баланс и завядание. Одним
из наиболее динамичных процессов в
растении является водный обмен, который
находится в тесной корреляций с другими
процессами жизнедеятельности растения.
Водный баланс — это поступление и
расходование воды растением. При
умеренной транспирации и достаточном
поступлении воды в растение создается
благоприятный водный баланс. В ясный
солнечный день это равновесие нарушается
и в растении возникает водный дефицит,
который обычно составляет 5—10%. Такой
дефицит считается вполне нормальным и
не приносит особого вреда растению.
При
интенсивной транспирации или иссушении
почвы, когда поступление воды в растение
прекращается, происходит значительная
потеря растительными клетками воды,
которая не пополняется поглощением ее
из почвы, в результате чего образуется
водный дефицит, часто наблюдаемый в
наиболее жаркие часы суток у растений.
При
водном дефиците листья теряют тургор,
завядают, повисают.
Некоторые
растения, имеющие в органах большое
количестве механических тканей, например
бессмертники (род. Helichrysum), не изменяют
своего внешнего вида при водном дефиците,
при значительной потере воды и даже при
гибели.
Наблюдения
показали, что обычно на рассвете
внутренний градиент в растении и почве
почти выравнивается, уравновешиваются
водные потенциалы растения и почвы. В
утренние часы, когда листья начинают
транспирировать, водный потенциал
становится несколько меньшим, чем на
рассвете, однако поступление воды в
растение начинается; когда создается
необходимый градиент водных потенциалов
от листьев к поверхности раздела
корень—почва.
Завядание
бывает временным и длительным.
Нарушение
водообмена. Его причины и последствия
Нарушение
водообмена у растений происходит при
недостаточном водоснабжении, то есть,
при наступлении засухи.
Засуха –
неблагоприятное сочетание метеорологических
условий, которые не обеспечивают
потребностей растения в воде. Засуха
бывает почвенная и атмосферная. Причины
почвенной засухи – отсутствие дождя
на протяжении длительного времени,
испарение с поверхности почвы и
транспирация, что приводит к высушиванию
корнеобитаемого слоя почвы. Последнюю
высушивают также сильные ветры. При
почвенной засухе влажность воздуха
бывает пониженной. Атмосферная засуха
вызывается массами сухого и нагретого
воздуха (суховея). Засуха, как почвенная,
так и атмосферная, задерживает рост
растений, уменьшает их листовую
поверхность, в результате чего урожай
снижается.
Влияние
почвенной (обезвоживание) и атмосферной
(перегрев) засух проявляется многими
признаками. Под влиянием обезвоживания
нарушается синтетическая способность
растений, наблюдается распад белков и
изменяется коллоидно-химическое
состояние цитоплазмы, снижается
количество накопленного растением
органического вещества из-за задержки
роста листьев и уменьшается их рабочая
поверхность. Особенно чувствительны к
засухе листья, находящиеся в фазе
эмбрионального роста. При подсыхании
из листьев исчезает крахмал. Усиленный
распад этого полисахарида не сопровождается
накоплением гексоз, поскольку они
используются в процессе дыхания. При
потере воды усиливается распад белковых
веществ.
Опыты
с растениями тыквы и огурца (В.Н. Жолкевич,
Т.Ф. Корецкая) показали, что почвенная
засуха вызывает глубокое нарушение
фосфорного обмена в корневой системе,
что приводит к уменьшению в корнях
количества АТФ, нуклеиновых кислот –
РНК и ДНК, уменьшению содержания белков
и увеличению в 7–10 раз сахаров, особенно
фруктозы и глюкозы. Задержка фосфорилирования
Сахаров вызывает уменьшение содержания
органических кислот – пировиноградной
кислоты, а также кислот цикла Кребса –
α-кетоглютаровой, янтарной, фумаровой,
щавелевоуксусной, яблочной и лимонной,
являющихся акцепторами аммиака при
синтезе аминокислот.
В
корнях и в пасоке повышается содержание
амидов, преимущественно глютамина.
Следовательно, симптомами угнетения
растений почвенной засухой прежде всего
является нарушение энергетического
обмена через уменьшение фосфорилирования,
а также задержку синтеза белков.
Длительные засухи приводят к резкому
снижению урожая.
Как
показали исследования Н.М. Сисакяна,
при обезвоживании растительных тканей
изменяется направленность действия
ферментов в сторону гидролиза и
задерживаются синтетические процессы
и рост. Ростовые процессы задерживаются
некоторое время даже после возобновления
водоснабжения, поскольку восстановление
синтетической направленности ферментов
происходит не сразу. Различные части
растения неодинаково устойчивы к засухе
и неодинаково на нее реагируют.
Установлено,
что растения, перенесшие небольшую
засуху, повторную переносят с меньшими
потерями. Однако устраивать искусственную
закалку взрослых растений в производственных
условиях трудно. П.А. Генкель предложил
подвергать закаливанию наклюнувшиеся
семена. Опыт показал, что растения,
выросшие из семян, подвергнутых
предпосевной закалке, приобретают
повышенную устойчивость к засухе.
Растворы
сахаров (глюкоза, галактоза, сахароза,
маннит, лактоза, мальтоза, раффиноза)
при инфильтрации в ткани листьев
значительно повышают их устойчивость
к высоким температурам. Считают, что
сахара «консервируют» структуру
митохондрии, которая становится
нечувствительной к тепловому влиянию,
и этим сохраняют функцию митохондрий
– процесс дыхания и окислительное
фосфорилирование (Ю.Г. Молотковский,
И.М. Жесткова).
Засухоустойчивые
сорта при значительном водном дефиците
характеризуются синтетической
направленностью в работе своих ферментов,
тогда как у менее засухоустойчивых
преобладает гидролитическая направленность.
Засухоустойчивые сорта озимой пшеницы
характеризуются большим содержанием
связанной воды, которая трудно обменивается
во время засухи, повышенной концентрацией
клеточного сока в период цветения и
налива зерна, высшим порогом коагуляции
белков, более интенсивным накоплением
сухого вещества зерна, более стойкой к
неблагоприятным условиям пигментной
системой. Этим и характеризуется
физиологическая природа засухоустойчивости
растений.
22
Основным
источником влаги является вода,
находящаяся в почве, и основным органом
поглощения воды — корневая система.
Роль этого органа, прежде всего,
заключается в том, что благодаря огромной
поверхности обеспечивается поступление
воды в растение из большего объема
почвы. Сформировавшаяся корневая система
представляет собой сложный орган с
хорошо дифференцированной внешней и
внутренней структурой.
Определение
размеров корневых систем требует
специальных методов. Очень много в этом
отношении достигнуто благодаря работам
русских физиологов В. Г. Ротмистрова,
А. П. Модестова, И. В. Красовской. Оказалось,
что общая поверхность корней обычно
превышает поверхность надземных органов
в 140-150 раз. Подсчитано, что число корней
у однолетних сеянцев яблони достигает
45 тыс. Корневые системы даже однолетних
хлебных злаков проникают в почву на
глубину 1,5-2 м. При выращивании одиночного
растения ржи было установлено, что общая
длина его корней может достигать 600км,
при этом на них образуется 15 млрд.
корневых волосков. Эти данные говорят
об огромной потенциальной способности
к росту корневых систем. Однако при
росте растений в фитоценозах, с достаточно
большой густотой их стояния, размеры
корневых систем заметно уменьшаются.
Рост
корня, его ветвление продолжаются в
течение всей жизни растительного
организма, т. е. практически он не
ограничен. Меристемы — образовательные
ткани — расположены на верхушке каждого
корня. Доля меристематических клеток
сравнительно велика (10% по массе против
1% у стебля).
Рост
корней отличается большой скоростью.
Считается, что одно растение риса в
благоприятных условиях может образовать
до 5 км новых корней в сутки. За счет
этого прироста корневой системы в
растение может дополнительно поступать
1,5 л воды. Только благодаря такому
интенсивному росту корневые системы
растения могут использовать скудно
рассеянную в почве воду. Важное значение
имеет явление гидротропизма, при котором
рост корневой системы как бы идет из
более иссушенных слоев почвы к более
влажным. В зависимости от типа растений
распределение корневой системы в почве
различно. У некоторых растений корневая
система проникает на большую глубину,
у других главным образом распространяется
в ширину.
24
По
степени доступности для растения
различают следующие формы почвенной
влаги. Гравитационная вода заполняет
промежутки между частицами почвы и
хорошо доступна растениям. Однако она
быстро испаряется и легко стекает в
нижние горизонты почвы под влиянием
силы тяжести, вследствие чего бывает в
почве лишь после дождей. Капиллярная
вода заполняет капилляры в почвенных
частицах. Эта вода хорошо доступна для
растений, она удерживается в капиллярах
силами поверхностного натяжения и
поэтому не только не стекает вниз, но и
поднимается вверх от грунтовых вод.
Пленочная вода окружает коллоидные
частицы почвы. Вода из периферических
слоев гидратационных оболочек может
поглощаться корнями. Гигроскопическая
вода адсорбируется сухой почвой при
помещении ее в атмосферу с 95 %-ной
относительной влажностью. Этот тонкий
слой молекул воды удерживается с такой
силой, что их водный потенциал достигает
-1000 бар и она недоступна для растений.
Количество
почвенной воды в процентах, при котором
растение впадает в устойчивое завядание,
называют коэффициентом или влажностью
завядания. Завядание растений разных
видов может начинаться при одной и той
же влажности, но промежуток времени от
завядания растения до его гибели
(интервал завядания) у растений может
быть различным. Так, для растений бобов
он составляет несколько суток, а для
растений проса – несколько недель.
Завядание начинается позже у растений
с более отрицательным осмотическим
потенциалом и меньшей скоростью
транспирации. «Мертвый запас» влаги в
почве – это количество воды полностью
недоступной растению. Он зависит от
механического состава почвы. Чем больше
глинистых частиц в почве, тем больше
«мертвый запас» влаги. Количество
доступной для растения воды представляет
собой разность между полевой влагоемкостью
(максимальное количество воды, удерживаемое
почвой) и «мертвым запасом».
Определение водного потенциала
Водный потенциал – это потенциальная энергия воды в системе по сравнению с чистой водой, когда температура и давление поддерживаются одинаковыми. Это также может быть описано как мера того, насколько свободно молекулы воды могут перемещаться в конкретной среде или системе. Он измеряется в килопаскалях (кПа) и представлен греческой буквой Psi (Ψ). Потенциал воды никогда не бывает положительным, но имеет максимальное значение ноль, то есть значение чистой воды при атмосферном давлении. Когда дело доходит до нечистой воды или воды, в которой есть растворенные вещества, тем больше растворенное вещество тем более отрицательным становится since, поскольку растворенные молекулы будут притягивать молекулы воды и ограничивать их свободу передвижения.
Движение молекул воды
Вода движется из областей, где водный потенциал выше (или менее отрицательный), в области, где он ниже (или более отрицательный), и мы называем это движение как осмос, Например, на диаграмме ниже решение вокруг клетка является гипертонический Это означает, что он имеет более высокую концентрацию растворенного вещества, поэтому более низкий потенциал воды, чем внутри клетки. Поскольку ячейка имеет частично проницаемую мембрану, позволяющую движение воды внутрь и наружу, вода будет перемещаться изнутри ячейки, где higher выше, во внешнюю часть ячейки, где Ψ ниже. Это может привести к гибели клеток в живых организмах. С другой стороны, клетка, помещенная в чистый водный раствор, может поглощать воду до тех пор, пока она не лопнет и не умрет. Следовательно, клетки нуждаются в среде, которая не отличается значительно по концентрации растворенного вещества.

Формула водного потенциала
Формула, используемая для расчета Ψ, следующая:
Ψ = Ψs + Ψp + Ψg + Ψm
Тем не менее, это часто упрощается как эта формула, что также правильно:
Ψ = Ψs + Ψp
Здесь standss обозначает растворенный потенциал, Ψp – потенциал давления, Ψg – гравитационный потенциал и Ψm – матричный потенциал. Потенциал давления относится к физическому давлению, которое объекты или мембраны клеток оказывают на молекулы воды, и оно увеличивается с увеличением давления. Обратите внимание, что потенциал давления обычно поддерживается положительным в растительных клетках, чтобы они могли сохранять свою форму, позволяя растению оставаться твердым. Кроме того, матричный потенциал учитывает силы между молекулами воды и поверхностями или веществами, такими как почва или клеточные мембраны. Матричный потенциал всегда отрицателен и является более значительным в сухих системах, таких как почвы, потому что мы обнаруживаем, что частицы воды прочно связаны с частицами почвы. Как следует из названия, гравитационный потенциал – это способ, которым гравитация Земли влияет на свободу движения молекул воды. Наконец, потенциал растворенного вещества зависит от количества растворенного вещества в растворе и уменьшается с увеличением концентрации растворенного вещества.
- диффузия – Произвольное распространение частиц из области, где они находятся в более высокой концентрации, в другую, где они находятся в более низкой концентрации.
- Гипотонический раствор – Раствор с более высокой концентрацией растворенного вещества по сравнению с другим раствором.
- Изотонический раствор – Раствор с концентрацией раствора, равной концентрации другого раствора.
- вязкость – Степень сопротивления жидкости.
викторина
1. Что мы можем ожидать, если поместить ячейку в раствор, где Ψ ячейки равно -0,3 кПа, а значение раствора -0,9 кПа?A. Вода выйдет из клеткиB. Вода переместится в клеткуC. Вода не будет двигаться в или из клеткиD. Клетка взорвется
Ответ на вопрос № 1
верно. Вода движется из области, где водный потенциал выше, в область, где она ниже. Это означает, что он переместится из клетки в раствор снаружи.
2. Проще говоря, водный потенциал:A. Количество воды, которое корни могут принимать в деньB. Сочетание осмотическое давление и гравитационные силыC. Сочетание растворенного потенциала и потенциала давленияD. Количество давления, необходимое для выталкивания воды вверх по стеблю растения
Ответ на вопрос № 2
С верно. Простая формула, которую мы используем: Ψ = растворенный потенциал (Ψs) + потенциал давления (Ψp).
3. Какое из следующих значений водного потенциала указывает на самую сухую окружающую среду?A. -0,1 пКаB. -1 рКаC. -0,03 пКаD. -5 пКа
Ответ на вопрос № 3
D верно. Чем суше окружающая среда, тем более она растворена и, следовательно, тем более отрицательно значение Ψ мы получаем.
4. Как меняется водный потенциал в зависимости от концентрации растворенного вещества?A. Это увеличивает, чем выше концентрация растворенного веществаB. Это уменьшается, чем выше концентрация растворенного веществаC. Это не зависит от концентрации растворенного веществаD. Концентрация растворенного вещества оказывает противоречивое влияние на него
Ответ на вопрос № 4
В верно. Чем выше концентрация растворенного вещества, тем менее свободны молекулы воды для перемещения и тем меньше is.

Содержание
- Компоненты водного потенциала
- Осмотический потенциал (Ψs)
- Матричный или матричный потенциал (m)
- Гравитационный или высотный потенциал (Ψg)
- Потенциал давления (Ψp)
- Методы определения водного потенциала
- Насос Шоландера или напорная камера
- Датчики давления
- Микрокапилляр с датчиком давления
- Вариации веса или объема
- Ожидаемые результаты и интерпретация
- Примеры
- Поглощение воды растениями
- Слизи
- Подъемный резервуар для воды
- Распространение воды в почве
- Ссылки
В водный потенциал Это свободная энергия или способность выполнять работу, имеющая определенный объем воды. Таким образом, вода на вершине водопада или водопада имеет высокий водный потенциал, который, например, способен приводить в движение турбину.
Символ, который используется для обозначения водного потенциала, – это заглавная греческая буква psi, которая пишется Ψ. Водный потенциал любой системы измеряется относительно водного потенциала чистой воды в условиях, считающихся стандартными (давление 1 атмосфера, высота и температура исследуемой системы).
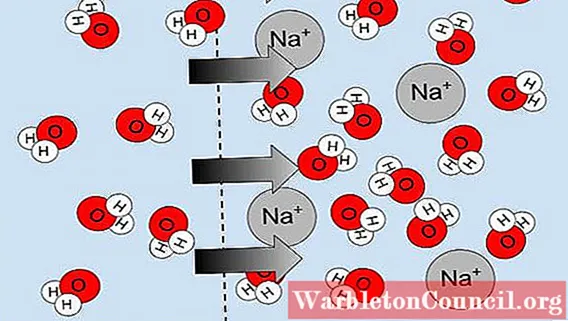
Факторами, определяющими водный потенциал, являются сила тяжести, температура, давление, гидратация и концентрация растворенных веществ, присутствующих в воде. Эти факторы определяют формирование градиентов водного потенциала, и эти градиенты управляют диффузией воды.
Таким образом, вода перемещается с участка с высоким потенциалом воды на другой с низким потенциалом воды. Компонентами водного потенциала являются осмотический потенциал (концентрация растворенных веществ в воде), матричный потенциал (адгезия воды к пористым матрицам), гравитационный потенциал и потенциал давления.
Знание водного потенциала необходимо для понимания функционирования различных гидрологических и биологических явлений. К ним относятся поглощение воды и питательных веществ растениями и поток воды в почве.
Компоненты водного потенциала
Водный потенциал состоит из четырех компонентов: осмотического потенциала, матричного потенциала, гравитационного потенциала и потенциала давления. Действие этих компонентов определяет наличие градиентов водного потенциала.
Осмотический потенциал (Ψs)
Обычно вода не находится в чистом виде, поскольку в ней растворены твердые вещества (растворенные вещества), например минеральные соли. Осмотический потенциал определяется концентрацией растворенных веществ в растворе.
Чем больше количество растворенных веществ, тем меньше свободной энергии воды, то есть меньше водный потенциал. Следовательно, вода пытается установить равновесие, перетекая из растворов с низкой концентрацией растворенных веществ в растворы с высокой концентрацией растворенных веществ.
Матричный или матричный потенциал (m)
В этом случае определяющим фактором является наличие матрицы или структуры гидратируемого материала, то есть он имеет сродство к воде. Это происходит из-за сил адгезии, создаваемых между молекулами, особенно водородных связей, образованных между молекулами воды, атомами кислорода и гидроксильными (ОН) группами.
Например, адгезия воды к почвенным глинам – это случай водного потенциала, основанного на матричном потенциале. Эти матрицы, притягивая воду, создают положительный водный потенциал, поэтому вода за пределами матрицы течет к ней и имеет тенденцию оставаться внутри, как это происходит в губке.
Гравитационный или высотный потенциал (Ψg)
В этом случае сила притяжения Земли определяет градиент потенциала, поскольку вода будет стремиться падать вниз. Вода, находящаяся на определенной высоте, имеет свободную энергию, определяемую притяжением, которое Земля оказывает на ее массу.

Например, вода в поднятом резервуаре для воды свободно падает по трубе и перемещается с этой кинетической энергией (движением), пока не достигнет крана.
Потенциал давления (Ψp)
В этом случае вода под давлением имеет большую свободную энергию, то есть больший водный потенциал. Следовательно, эта вода будет перемещаться из того места, где она находится под давлением, туда, где его нет, и, следовательно, будет меньше свободной энергии (меньший водный потенциал).
Например, когда мы дозируем капли с помощью пипетки, когда мы нажимаем резиновую ручку, мы оказываем давление, которое дает энергию воде. Из-за этой более высокой свободной энергии вода движется наружу, где давление ниже.
Методы определения водного потенциала
Существует множество методов измерения водного потенциала, некоторые из которых подходят для почвы, другие – для тканей, для механических гидравлических систем и другие. Водный потенциал эквивалентен единицам давления и измеряется в атмосферах, барах, паскалях или фунтах на квадратный дюйм (фунты на квадратный дюйм в английской аббревиатуре).
Вот некоторые из этих методов:
Насос Шоландера или напорная камера
Если вы хотите измерить водный потенциал листа растения, вы можете использовать напорную камеру или насос Шоландера. Он состоит из герметичной камеры, в которую помещается весь лист (лист с черешком).
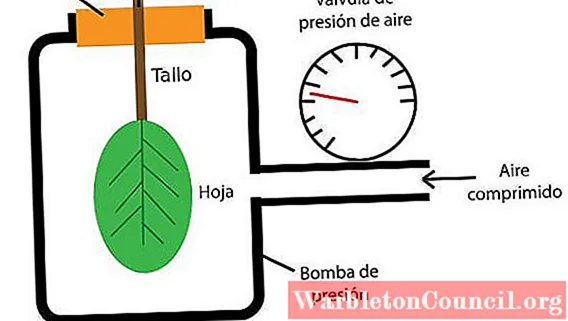
Затем давление внутри камеры повышают за счет подачи сжатого газа, измеряя давление, которое достигается с помощью манометра. Давление газа на лист увеличивается до такой степени, что содержащаяся в нем вода вырывается через сосудистую ткань черешка.
Давление, показываемое манометром, когда вода покидает лист, соответствует водному потенциалу листа.
Датчики давления
Есть несколько альтернатив для измерения водного потенциала с помощью специальных инструментов, называемых датчиками давления. Они предназначены для измерения водного потенциала почвы, основанного в основном на матричном потенциале.
Например, есть цифровые датчики, которые работают на основе введения в почву пористой керамической матрицы, подключенной к датчику влажности. Эта керамика увлажняется водой внутри почвы до тех пор, пока не будет достигнут баланс между водным потенциалом в керамической матрице и водным потенциалом почвы.
Затем датчик определяет влажность керамики и оценивает водный потенциал почвы.
Микрокапилляр с датчиком давления
Существуют также зонды, способные измерять водный потенциал в тканях растений, таких как стебель растения. Одна модель состоит из очень тонкой трубки с тонким наконечником (микропиллярной трубки), которая вводится в ткань.
Проникая в живую ткань, раствор, содержащийся в клетках, следует градиенту потенциала, определяемому давлением, содержащимся в стержне, и вводится в микропиле. Когда жидкость из штока попадает в трубку, она выталкивает содержащееся в ней масло, которое активирует датчик давления или манометр, который устанавливает значение, соответствующее потенциалу воды.
Вариации веса или объема
Чтобы измерить водный потенциал на основе осмотического потенциала, можно определить вариации веса ткани, погруженной в растворы с различными концентрациями растворенного вещества. Для этого готовят серию пробирок, каждая с известной возрастающей концентрацией растворенного вещества, например сахарозы (сахара).
То есть, если в каждой из 5 пробирок по 10 см3 воды, в первую пробирку добавляют 1 мг сахарозы, во вторую – 2 мг и, таким образом, до 5 мг в последнюю. Итак, у нас есть возрастающая батарея концентраций сахарозы.
Затем из ткани, водный потенциал которой необходимо определить, вырезают 5 кусков равного известного веса (например, кусочки картофеля). Затем в каждую пробирку помещают срез, и через 2 часа срезы ткани удаляют и взвешивают.
Ожидаемые результаты и интерпретация
Ожидается, что одни кусочки потеряют вес из-за потери воды, другие наберут его, потому что впитали воду, а третьи сохранят вес.
Те, кто потеряли воду, находились в растворе, в котором концентрация сахарозы была больше, чем концентрация растворенных веществ в ткани. Следовательно, вода текла в соответствии с градиентом осмотического потенциала от самой высокой концентрации до самой низкой, и ткань теряла воду и вес.
Напротив, ткань, которая набрала воду и вес, была в растворе с более низкой концентрацией сахарозы, чем концентрация растворенных веществ в ткани. В этом случае градиент осмотического потенциала благоприятствовал проникновению воды в ткань.
Наконец, в том случае, когда ткань сохранила свой первоначальный вес, делается вывод, что концентрация, в которой она была обнаружена, имеет такую же концентрацию растворенного вещества. Следовательно, эта концентрация будет соответствовать водному потенциалу исследуемой ткани.
Примеры
Поглощение воды растениями
Дерево высотой 30 м должно транспортировать воду от земли до последнего листа, и это осуществляется через его сосудистую систему. Эта система представляет собой специализированную ткань, состоящую из мертвых клеток, которые выглядят как очень тонкие трубки.

Перенос возможен благодаря разнице в водном потенциале, который создается между атмосферой и листом, который, в свою очередь, передается в сосудистую систему. Лист теряет воду в газообразном состоянии из-за более высокой концентрации в нем водяного пара (более высокий водный потенциал) по сравнению с окружающей средой (более низкий водный потенциал).
Потеря пара создает отрицательное давление или всасывание, которое направляет воду из сосудов сосудистой системы к листовой пластинке. Это всасывание передается от сосуда к сосуду до тех пор, пока не достигнет корня, где клетки и межклеточные пространства впитываются водой, поглощенной из почвы.
Вода из почвы проникает в корень из-за разницы в осмотическом потенциале воды в клетках эпидермиса корня и почвы. Это происходит потому, что в клетках корней растворенные вещества выше, чем в почвенной воде.
Слизи
Многие растения в засушливой среде задерживают воду, производя слизь (вязкое вещество), которая хранится в их вакуолях. Эти молекулы удерживают воду, уменьшая их свободную энергию (низкий водный потенциал), в этом случае решающим является матричный компонент водного потенциала.
Подъемный резервуар для воды
В случае системы водоснабжения на основе приподнятого резервуара он заполняется водой из-за действия потенциала давления.Компания, предоставляющая услуги водоснабжения, оказывает давление на нее с помощью гидравлических насосов и, таким образом, преодолевает силу тяжести, чтобы достичь резервуара.
Когда бак наполняется, вода распределяется из него благодаря разнице потенциалов между водой, хранящейся в баке, и выходами воды в доме. Открытие крана устанавливает градиент гравитационного потенциала между водой в кране и водой в баке.
Следовательно, вода в резервуаре имеет больше свободной энергии (более высокий водный потенциал) и падает в основном под действием силы тяжести.
Распространение воды в почве
Основным компонентом водного потенциала почвы является матричный потенциал, учитывая силу сцепления, которая устанавливается между глинами и водой. С другой стороны, потенциал силы тяжести влияет на градиент вертикального смещения воды в почве.
Многие процессы, происходящие в почве, зависят от свободной энергии воды, содержащейся в почве, то есть от ее водного потенциала. Эти процессы включают питание растений и транспирацию, проникновение дождевой воды и испарение воды из почвы.
В сельском хозяйстве важно определить водный потенциал почвы для правильного полива и удобрения. Если матричный потенциал почвы очень высок, вода будет оставаться связанной с глинами и не будет доступна для поглощения растениями.
Ссылки
- Буссо, К.А. (2008). Использование барокамеры и термопарных психрометров для определения водных отношений в тканях растений. ФИТОН.
- Квинталь-Ортис, В.К., Перес-Гутьеррес, А., Латурнери-Морено, Л., Май-Лара, К., Руис-Санчес, Э. и Мартинес-Чакон, А.Дж. (2012). Использование воды, водный потенциал и урожайность перца хабанеро (Capsicum chinense Jacq.). Журнал Fitotecnia Mexicana.
- Солсбери, Ф. и Росс, К. (1991). Физиология растений. Wadsworth Publishing.
- Scholander, P., Bradstreet, E., Hemmingsen, E. и Hammel, H. (1965). Давление сока в сосудистых растениях: у растений можно измерить отрицательное гидростатическое давление. Наука.
- Squeo, F.A. (2007). Вода и водный потенциал. В: Squeo, F.A. и Cardemil, L. (ред.). Физиология растений. Издания Университета Ла Серены
