К классу Пресмыкающиеся, или Рептилии, относится около (10) тыс. видов наземных животных, которые в отличие от Земноводных не имеют тесной связи с водой. К ним относятся ящерицы, змеи, крокодилы, черепахи и другие животные.
Внешнее строение пресмыкающихся

Рис. (1). Внешнее строение ящерицы
Тело пресмыкающихся разделяется на голову, туловище, хвост, передние и задние конечности (отсутствуют у змей). Конечности располагаются по бокам туловища и прижимаются к земле (как бы смыкаются с поверхностью), поэтому этот класс получил такое название — пресмыкающиеся.
Кожа сухая, не содержит желез. Кожные покровы образованы ороговевшим эпидермисом (роговыми чешуйками, или щитками). Роговые чешуйки предохраняют животное от испарения влаги и иссушения.

Рис. (2). Роговой покров
Растут пресмыкающиеся во время периодических линек.

Рис. (3). Линька
Происхождение
Первые рептилии появились на планете около (280) миллионов лет назад.
Их предками считают стегоцефалов — древних земноводных.

Рис. (4). Стегоцефал
Развитие и расцвет рептилий связаны с изменением климатических условий: в мезозойской эре он становится более сухим, уменьшается количество заболоченных площадей.
В новых условиях преимущество получили те земноводные, которые не имели тесной связи с водой. От таких древних земноводных и произошли первые пресмыкающиеся.
Новые особенности, появившиеся у рептилий (по сравнению с земноводными), перечислены ниже.
Обрати внимание!
- Сухая кожа, без желез, покрытая роговыми чешуйками;
- в желудочке сердца появилась неполная перегородка, артериальная и венозная кровь полностью не смешиваются;
- более совершенными стали органы дыхания — появились дыхательные пути, увеличилась внутренняя поверхность лёгких благодаря ячеистому строению;
- развитие зародыша происходит в яйце, он защищён оболочками;
- оплодотворение внутреннее, оно происходит в половых органах самки;
- развитие без личиночной стадии (прямое);
- более сложным стал мозг — увеличился передний отдел, появились зачатки коры.
Расцвет пресмыкающихся наступил (225) миллионов лет назад. Они заселили все среды обитания. В воде плавали плезиозавры — «морские ящеры» и ихтиозавры — «рыбоящеры». Среди наземных животных царствовали разнообразные динозавры — «ужасные ящеры». В воздухе парили птерозавры — «летающие ящеры».

Рис. (5). Птерозавры
Самые древние рептилии, дожившие до наших дней, — крокодилы и черепахи.

Рис. (6). Крокодилы
Одна из групп ящеров — Зверозубые ящеры — считается переходной формой между пресмыкающимися и млекопитающими. Самым знаменитым зверозубым ящером является Иностранцевия.

Рис. (7). Зверозубый ящер
- Уничтожают насекомых и грызунов;
- некоторые используются в пищу человеком;
- из кожи крокодилов и змей, панцири черепах изготавливают разные предметы;
- змеиный яд находит применение в фармакологии.
Источники:
Рис. 1. Внешнее строение ящерицы. https://image.shutterstock.com/image-photo/one-green-lizard-isolated-on-600w-1683855673
Рис. 2. Роговой покров. https://cdn.pixabay.com/photo/2016/06/17/21/44/snake-1464184. 22.11.2021.
Рис. 3. Линька. https://image.shutterstock.com/image-photo/snake-molt-on-ground-600w-1919916737
Рис. 4. Стегоцефал. https://image.shutterstock.com/image-illustration/realistic-drawing-ichthyostega-animals-paleozoic-600w-1699920664
Рис. 5. Птерозавры. https://image.shutterstock.com/image-illustration/flying-pterodactyl-over-land-3d-600w-513495175
Рис. 6. Крокодилы. https://cdn.pixabay.com/photo/2015/03/16/22/25/crocodiles-676946. 22.11.2021.
Рис. 7. Зверозубый ящер. https://www.shutterstock.com/ru/image-illustration/inostrancevia-3d-illustration-1128076058
Пресмыкающиеся
по сравнению с земноводными представляют
собой следующий этап приспособления
позвоночных животных к жизни на суше.
Это первый настоящий
класс наземных позвоночных животных.
Обитают они преимущественно в
регионах с теплым и жарким климатом. В
ходе завоевания суши пресмыкающиеся
приобрели ряд адаптации:
-
Тело
подразделяется на голову, шею, туловище,
хвост и пятипалые
конечности. -
Кожа
сухая, лишена желез и покрыта роговым
покровом,
защищающим
тело от высыхания. Рост животного
сопровождается
периодической линькой. -
Скелет
прочный,
окостеневший. Позвоночник
состоит из пяти отделов:
шейного, грудного, поясничного,
крестцового и хвостового.
Плечевой и тазовый пояса конечностей
укреплены и
связаны с осевым скелетом.
Развиты ребра и грудная клетка. -
Мускулатура
более дифференцирована, чем у земноводных.
Развиты
шейные
и межреберные мышцы, подкожная
мускулатура. Движения
отделов тела более разнообразные и
быстрые. -
Пищеварительный
тракт более длинный, чем у земноводных,
и четче
дифференцирован на отделы. Пища
захватывается челюстями,
имеющими
многочисленные острые зубы. Стенки
ротовой полости и пищевода снабжены
мощной мускулатурой, которая
проталкивает крупные порции пищи в
желудок. На
границе
тонкой и толстой кишок имеется слепая
кишка, особенно
хорошо развитая у растительноядных
наземных черепах. -
Органы
дыхания – легкие
– имеют
большую дыхательную
поверхность
за счет ячеистого строения. Развиты
воздухопроводящие
пути – трахея,
бронхи, в
которых воздух увлажняется
и не иссушает легкие. Вентиляция легких
происходит путем изменения
объема грудной клетки.
-
Сердце
трехкамерное,
однако
в желудочке имеется неполная
продольная
перегородка, препятствующая полному
смешиванию
артериальной и венозной крови. Большая
часть тела
пресмыкающихся
снабжается смешанной кровью с
преобладанием артериальной, поэтому
интенсивность обмена выше, чем
у земноводных. Однако пресмыкающиеся,
так же как рыбы и
земноводные, являются пойкилотермными
(холоднокровными)
животными, температура
тела которых зависит от температуры
среды обитания. -
Органы
выделения – тазовые
почки. По
мочеточникам моча
оттекает
в клоаку, а из нее – в мочевой пузырь.
В нем вода
дополнительно
отсасывается в кровеносные капилляры
и возвращается
в организм, после чего моча выводится
наружу.
Конечный
продукт азотистого обмена, выводимый
с мочой, – мочевая
кислота. -
Головной
мозг имеет больший относительный
размер, чем у
земноводных.
Лучше развиты большие полушария
переднего
мозга
с зачатками коры
и
мозжечок. Формы поведения пресмыкающихся
более сложные. Органы чувств лучше
приспособлены
к наземному образу жизни.
10.
Оплодотворение только внутреннее.
Яйца,
защищенные от высыхания
кожистой или скорлуповой оболочкой,
рептилии откладывают
на
суше. Зародыш
в яйце развивается в водной оболочке.
Развитие прямое.
Особенности
строения и процессов жизнедеятельности.
Рассмотрим
строение основных органов пресмыкающихся
на примере
ящерицы
прыткой.
Тело
ящерицы подразделено на голову, туловище
и хвост. В
туловищном отделе хорошо выражена шея.
Все тело покрыто роговыми
чешуями, а голова и брюхо — крупными
щитками. Конечности
ящерицы хорошо развиты и вооружены
пятью пальцами с когтями.
Плечевые
и бедренные кости расположены параллельно
поверхности
земли, вследствие чего тело провисает
и касается земли (отсюда
и название класса). Шейный отдел
позвоночника состоит из восьми
позвонков, первый из них подвижно
соединён как с черепом, так и
со вторым позвонком, что обеспечивает
головному отделу большую
свободу движений. Позвонки пояснично-грудного
отдела несут
ребра, часть которых соединена с грудиной,
в результате чего образуется
грудная клетка. Крестцовые позвонки
обеспечивают более
прочное, чем у земноводных, соединение
с костями таза.
У
ящериц при самопроизвольном отбрасывании
хвоста (явление
автотомии) разрыв происходит не между
позвонками, а посередине,
где находятся тонкие хрящевые прослойки,
делящие тело
позвонка на две части.
Органы
выделения представлены
тазовыми почками, в которых общая
фильтрационная площадь клубочков
небольшая, в то время как
протяженность канальцев значительная.
Это способствует интенсивному
обратному всасыванию отфильтрованной
клубочками воды
в капилляры крови. Следовательно,
выделение продуктов жизнедеятельности
у пресмыкающихся происходит с минимальными
потерями
воды. У них, как и у наземных членистоногих,
конечным продуктом
выделения является мочевая кислота,
требующая для выведения
из организма небольшого количества
воды. Моча по мочеточникам
собирается в клоаку, а из нее в мочевой
пузырь, из которого
в виде взвеси мелких кристаллов выводится
наружу.
Головной
мозг пресмыкающихся
по сравнению с таковым земноводных
обладает лучше развитыми мозжечком и
большими полушариями переднего
мозга, поверхность которого имеет
зачатки
коры. Это обусловливает разнообразные
и более сложные формы приспособительного
поведения.
Органы
чувств в
большей мере соответствуют наземному
образу
жизни. Глаза защищены подвижными веками
(верхним и нижним)
и мигательной перепонкой. Фокусировка
зрения достигается
как перемещением хрусталика относительно
сетчатой оболочки,
так и изменением его кривизны. Некоторые
дневные виды обладают цветовым зрением.
У ящериц хорошо развит теменной глаз
– светочувствительный орган, расположенный
на темени.
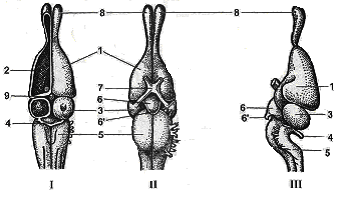
Рис.
41. Головной мозг ящерицы: I
–
сверху; II
– снизу; III
– сбоку; 1 – передний мозг; 2 – полосатое
тело; 3 – средний мозг; 4 –
мозжечок; 5 – продолговатый мозг; 6 –
воронка; 6′ – гипофиз;
7 – хиазма; 8 – обонятельная доля; 9 –
эпифиз.
Орган
слуха состоит
из среднего и внутреннего уха. Обоняние
по сравнению с земноводными развито
лучше.
Некоторые
виды змей имеют орган термического
чувства (между
ноздрями и глазом), позволяющий на
расстоянии улавливать тепло, исходящее
от объекта добычи. Это дает возможность
змеям охотиться на теплокровных животных,
не видя их.
У
пресмыкающихся оплодотворение внутреннее.
Размножаются
путем откладки яиц или яйцеживорождения.
Яйца относительно крупные,
богаты питательными веществами, что
обеспечивает прямое
развитие эмбриона без промежуточных
личиночных стадий. Яйца снаружи защищены
от высыхания защитными оболочками
(кожистой или скорлуповой). Зародыш в
яйце развивается в полости,
заполненной жидкостью, что способствует
правильному формированию
его органов.
Разнообразие
и значение пресмыкающихся
Современные
пресмыкающиеся представляют собой
лишь небольшие остатки богатого
и разнообразного мира животных, населявших
в мезозойскую
эру не только всю сушу, но и все моря
планеты. В настоящее время
к классу Пресмыкающиеся принадлежат
более 7 тыс. видов,
объединенных в несколько отрядов, среди
которых наиболее многочисленными
являются Чешуйчатые, Крокодилы, Черепахи
и Клювоголовые.
Отряд
Чешуйчатые
(Sguamata)
–
самая многочисленная группа пресмыкающихся
(примерно 6,5 тыс. видов). Для них характерно
наличие
в покровах роговых чешуи.
В
средней полосе СНГ обитает ящерица
прыткая, севернее распространена
ящерица живородящая, а в южных районах
обитают
гекконы, агамы и самая крупная ящерица
– серый варан (до
2 м длиной). Варан благодаря хорошо
развитым конечностям быстро
бегает, его тело высоко поднято над
землей. Распространены
вараны в Африке, Южной Азии, Малайском
архипелаге и в Австралии, а также в
песчаных пустынях Туркменистана и
Узбекистана.
Змеи
— это безногие чешуйчатые с длинным
цилиндрическим
телом, с помощью волнообразных изгибов
которого они передвигаются.
Подвижных век не имеют. Добычу заглатывают
целиком
благодаря широко растяжимому рту (нижние
челюсти подвешены
на растяжимых связках). Зубы острые,
направлены назад.
При нападении на жертву ядовитые змеи
выдвигают вперед
из ротовой полости зубы и с их помощью
вводят в тело добычи секрет ядовитых
желез. Грудина отсутствует. Ребра
свободные и
исключительно подвижные. Среднее ухо
упрощено, барабанная
перепонка отсутствует. Распространены
во всех частях света, но
численно преобладают в жарких странах.
Широко известны неядовитые
змеи – ужи, удавы, и ядовитые – гюрза,
гадюка, гремучая
змея, песчаная эфа и др. Яд змей используется
для приготовления
лекарственных препаратов.
Отряд
Крокодилы (Crocodylia)
представлен
крупными (длиной до 6 м), наиболее
высокоорганизованными пресмыкающимися,
приспособленными
к полуводному образу жизни. У них
ящерообразное,
слегка уплощенное тело, покрытое роговыми
щитками, со сжатым с боков хвостом и
плавательными перепонками между пальцами
задних ног.
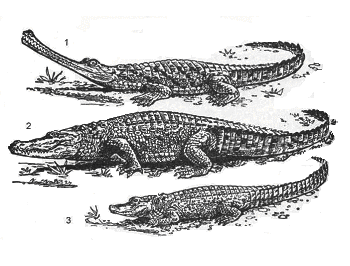
Рис.
42. Крокодилы: 1
– гавиал; 2 – нильский крокодил; 3 – китайский
аллигатор
Зубы
сидят в ячейках (как у млекопитающих).
Основания зубов внутри полые, в этих
полостях развиваются новые, замещающие
зубы. Смена зубов в течение жизни
крокодила многократна. Легкие имеют
сложноячеистое строение и вмещают
большой
запас воздуха. Развита диафрагма. Сердце
четырехкамерное.
Размножаются
откладкой яиц (10—100 штук), покрытых
известковой
скорлупой. Становятся половозрелыми к
8—10 годам, живут до
80—100 лет.
Известны
нильский крокодил (Африка), аллигатор
(Китай,
Америка), кайман (Америка), гавиал
(Индостан, Бирма), В
некоторых странах мясо крокодилов
используется в пишу, кожа является
ценным сырьем для изготовления
галантерейных изделий. В
связи с интенсивным промыслом численность
крокодилов резко сократилась. Созданы
хозяйства по их разведению (США, Куба).
Отряд
Черепахи (Testudines)
объединяет
пресмыкающихся, имеющих компактное
тело, заключенное в прочный костный
панцирь, в который
могут втягиваться шея, голова, конечности
и хвост. Сверху костный
панцирь покрыт роговыми пластинками
или мягкой кожей.

Рис.
43. Черепахи: 1
– слоновая черепаха; 2 – степная черепаха;
3 – болотная черепаха; 4
– каретта; 5 – уссурийская мягкокожистая
черепаха.
Челюсти
лишены зубов и имеют острые роговые
края. Позвонки,
кроме шейного и хвостового отделов,
сращены со спинной
частью панциря (как и ребра). Механизм
дыхания связан с движением
шеи и плеч, которые, выдвигаясь из-под
панциря, растягивают легкие. Интенсивность
обмена низкая. Способны к длительному
голоданию. Живут во влажных тропиках и
в жарких пустынях.
Во многих странах мясо и яйца черепах
употребляют в пищу.
Роговые пластины некоторых видов черепах
используются для
изготовления поделок. Болотная
черепаха – живет
в слабо проточных водоемах
и питается разнообразными мелкими
водными и наземными животными.
На
Галапагосских островах живёт слоновая
черепаха. Громадный
панцирь бывает длиной до 110 см и высотой
до 60 см. толстые и мощные столбообразные
ноги поддерживают тяжеловесное туловище.
Масса взрослых экземпляров составляет
около 100 кг, а отдельных гигантов – до
400 кг.
Единственный
вид современных Клювоголовых
(Rhynchocephalia)
–
гаттерия
имеет много чрезвычайно примитивных
черт и сохранилась только в Новой
Зеландии и на прилежащих островах.

Рис.
44. Гаттерия.
Гаттерия
внешне очень напоминает ящерицу с
массивным телом, большой головой и
пятипалыми конечностями. От затылка по
спине и хвосту тянется невысокий гребень
из треугольных вертикальных пластинок.
Окрашена гаттерия в тусклый оливково-зелёный
цвет, на боках тела и конечностях мелкие
и более крупные желтые пятна.
Зрачки
больших глаз, расположенных по бокам
головы, в виде вертикальной щели.
Барабанных перепонок у гаттерии нет,
полость среднего уха заполнена жировой
тканью.
Тело
взрослых самцов длиной до 60 см, массой
800 г. самки почти вдвое мельче самцов.
Гаттерия достигает половозрелости лишь
к 20 годам. Велика и продолжительность
жизни: в неволе гаттерии жили более 70
лет.
Основная
пища гаттерии – различные беспозвоночные,
особенно насекомые, в частности жуки и
крупные бескрылые кузнечики, а также
пауки, черви, моллюски, иногда ящерицы,
лягушки, яйца птиц. Добычу гаттерия
проглатывает целиком.
Передвигается
гаттерия медленно, при этом почти не
поднимает брюхо над субстратом. Однако
при охоте или в испуганном состоянии
она приподнимается на ногах и двигается
быстро. Кроме того, она хорошо плавает
и охотно заходит в воду.
Происхождение
пресмыкающихся.
Пресмыкающиеся
известны
уже с конца каменноугольного периода
палеозойской эры. Своего
расцвета они достигли в мезозойскую
эру, к концу которой
были вытеснены птицами и млекопитающими.
Предками современных
пресмыкающихся считаются примитивные
девонские
земноводные – стегоцефалы, давшие
начало котилозаврам
– древним пресмыкающимся.
Расцвету
древних пресмыкающихся в мезозойскую
эру способствовали
теплый климат, обилие пищи как на суше,
так и в воде, а также отсутствие
конкурентов. Они заселили наземную
среду, где господствовали
гигантские динозавры, достигавшие в
длину 30 м. Среди них были как растительноядные
животные, так и хищники. В
водной среде господствовали рыбообразные
ящеры – ихтиозавры
(8 – 12 м). Своеобразную группу составляли
ящеры-птерозавры, которые могли
летать благодаря большой кожистой
перепонке, натянутой между передними
и задними конечностями.
Вымирание
древних пресмыкающихся связывают с
похолоданием
климата в конце мезозоя и неспособностью
их поддерживать
постоянную температуру тела. Наступившее
снижение процессов
жизнедеятельности у пресмыкающихся
привело к ослаблению
их конкурентной борьбы с появившимися
и быстро прогрессирующими
млекопитающими.
Контрольные
вопросы:
-
В
чём особенность организации пресмыкающихся? -
Какие
отряды входят в класс пресмыкающихся? -
Какие
особенности строения скелета присущи
пресмыкающимся? -
Что
такое автотомия у пресмыкающихся? -
Какие
особенности размножения характерны
пресмыкающимся?
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Эта статья о парафилетической группе; о кладе см. Завропсиды.
Запрос «Рептилия» перенаправляется сюда; см. также другие значения.
| Пресмыкающиеся | |||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||
|
Домен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Вторичноротые Тип: Хордовые Подтип: Позвоночные Инфратип: Челюстноротые Надкласс: Четвероногие Клада: Амниоты Клада: Завропсиды Класс: Пресмыкающиеся |
|||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||
| Reptilia Laurenti, 1768 | |||||||||||||||||||||||||||||||||||||
| Современные отряды | |||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||
| Геохронология
появился 312 млн лет
◄ Наше время ◄ Мел- ◄ Триасовое вымирание ◄ Массовое пермское вымирание ◄ Девонское вымирание ◄ Ордовикско-силурийское вымирание ◄ Кембрийский взрыв |
|||||||||||||||||||||||||||||||||||||
|


Пресмыка́ющиеся, или репти́лии (лат. Reptilia), — традиционно выделяемый класс преимущественно наземных позвоночных животных из клады амниот, включающий современных черепах, крокодилов, клювоголовых и чешуйчатых. Пресмыкающиеся являются парафилетической группой, так как дали начало птицам, но не включают их самих. В качестве монофилетической альтернативы класса выделяется клада завропсид, или рептилий в пересмотренном составе, в которую включают представителей традиционного класса пресмыкающихся (кроме зверообразных) и птиц[1][2][3][4].
В XVIII—XIX веках вместе с амфибиями объединялись в группу гадов — холоднокровных наземных позвоночных.
Традиционно к этой группе относили различных позвоночных, согласно изначальным представлениям похожих по своей организации на современных рептилий (например, синапсид — предков и родственников современных млекопитающих). Однако в настоящее время ряд вопросов о физиологии многих вымерших групп организмов остаётся открытым, а данные об их генетических и эволюционных связях не поддерживают подобного рода классификацию.
Рептилии пережили расцвет в эпоху мезозоя, когда они доминировали на суше, в море и в воздухе. Крупнейшие наземные животные принадлежали к динозаврам. В конце мелового периода большая часть рептилий, особенно крупного размерного класса, вымерла. Однако бытующее мнение, что современные пресмыкающиеся являются лишь разрозненными остатками того древнего мира, неверно. Выделяется около 11 940 видов пресмыкающихся, большая часть которых относится к отряду чешуйчатых[5], на территории России обитает 77 видов[6][7][обновить данные]. Небольшие хищные динозавры дали начало процветающей в настоящее время группе животных — птицам, причём многие приспособления, обусловившие эволюционный успех этой группы, появились ещё у её архозавровых предков, представлявших собой специализированную группу диапсид (теплокровность, теплоизолирующий покров тела — перья, развитый головной мозг, и так далее).
Эволюция пресмыкающихся[править | править код]
Предки пресмыкающихся — рептилиоморфы — известны со среднего карбона. Ранее первыми рептилиями считали представителей группы котилозавров, в которую включали различные малосвязанные таксоны рептилиоморф. Продвинутые рептилиоморфы — амниоты — сразу разделились на две ветви: синапсиды и завропсиды. К синапсидам относятся млекопитающие и их вымершие родственники, ранее известные как «зверообразные рептилии» (с точки зрения кладистической таксономии пресмыкающимися не являются). К завропсидам, в свою очередь, относятся пресмыкающиеся и филогенетически вложенные в их состав птицы. В конце пермского периода оформляются основные эволюционные линии рептилий, включая архозавроморф и лепидозавроморф. В триасе появляются первые динозавры, крокодиломорфы, завроптеригии, первые черепахи и прочие группы. В мезозойской эре наступает расцвет пресмыкающихся, среди представителей наблюдается наибольшее многообразие. Происходит освоение морских и речных водоёмов, а также воздушного пространства. В юрском периоде одна из ветвей тероподовых динозавров — манирапторы — эволюционирует в первых птиц (авиал). Последняя группа — змеи — сформировалась в меловом периоде.
В конце мелового периода происходит резкое сокращение количества видов пресмыкающихся. Однозначно указать причины вымирания современная наука пока не может[источник не указан 410 дней].
Происхождение[править | править код]
Произошли рептилии примерно 310—320 миллионов лет назад, в болотах позднего каменноугольного периода, когда из продвинутых рептиломорфов стали выделяться другие виды[8].
Самым старым известным животным, которое могло быть амниотой, является Casineria (хотя, возможно, это был темноспондил). Обнаружена целая серия следов из ископаемых пластов Новой Шотландии, датированных 315 миллионов лет назад; видны типичные для рептилий пальцы ног и отпечатки чешуи[9]. Эти следы приписываются Hylonomus, старейшей из известных рептилий[10]. Это было маленькое, похожее на ящерицу животное, от 20 до 30 сантиметров (от 7,9 до 11,8 дюйма) в длину, с многочисленными острыми зубами, указывающими на питание в основном насекомыми[11]. Другие примеры включают Westlothiana (на данный момент считается лишь рептилиоморфом, а не настоящей амниотой) и Paleothyris, оба похожего телосложения[12].
Однако иногда микрозавров считали настоящими рептилиями, поэтому возможно более раннее происхождение пресмыкающихся[12].
Дальнейшая история[править | править код]
Самые ранние амниоты, в том числе первые рептилии (те амниоты, которые ближе к современным рептилиям, чем к млекопитающим), были в значительной степени затенены более крупными четвероногими, такими как Cochleosaurus, и оставались небольшой, незаметной частью фауны до краха каменноугольных тропических лесов. Этот внезапный коллапс затронул несколько больших групп. Примитивные четвероногие были особенно затронуты, многие исчезли, в то время как рептилии жили лучше, будучи экологически адаптированными к более засушливым условиям. Примитивным четвероногим, как и современным амфибиям, было необходимо вернуться в воду, чтобы отложить яйца; напротив, амниоты, как и современные рептилии, яйца которых имеют скорлупу, позволяющую откладывать их на сушу, лучше приспособились к новым условиям жизни. Амниоты занимали новые ниши быстрее, чем до коллапса, и гораздо быстрее, чем примитивные четвероногие. Они приобрели новые стратегии питания, в том числе травоядные и плотоядные (ранее они были только насекомоядными и рыбоядными). С этого момента рептилии доминировали в пищевых цепях и отличались большим разнообразием, чем примитивные четвероногие, что подготовило почву для мезозоя (известного как эпоха рептилий). Одной из самых известных ранних рептилий является Mesosaurus, род из ранней перми, который обитал в воде, питаясь рыбой[13][14].
Исследование разнообразия рептилий в каменноугольном и пермском периодах, проведённое в 2021 году, предполагает гораздо более высокую степень разнообразия, чем считалось ранее, сопоставимую или даже превышающую таковую у синапсид. Таким образом началась «Первая эпоха рептилий»[12].
Анапсиды, синапсиды, диапсиды и завропсиды[править | править код]
Традиционно предполагалось, что первые рептилии сохранили унаследованный от предков анапсидный череп, имеющий отверстия только для глаз и ноздрей. Открытие синапсидоподобных отверстий (см. ниже) в черепе нескольких представителей Parareptilia (клады, содержащей большинство амниот, традиционно называемых «анапсидами»), включая лантанозухоидов, миллереттид, болозавридов, некоторые никтеролетериды, некоторые проколофоноиды и, по крайней мере, некоторые мезозавры сделало его более двусмысленным, и в настоящее время неясно, имел ли предковый амниот анапсидоподобный или синапсидоподобный череп. Этих животных традиционно называют «анапсиды», и они образуют парафилетическую группу, из которого произошли другие группы. Очень скоро после того, как появились первые амниоты, отделились животные, названные Synapsida. Это «амниоты, похожие на млекопитающих», или предки млекопитающих, которые позже дали начало настоящим млекопитающим. Вскоре после этого другая группа развила аналогичную черту, на этот раз с двойным отверстием за каждым глазом, за что они получили название Diapsida («две арки»). Функция отверстий в этих группах заключалась в том, чтобы облегчить череп и дать место для движения мышц челюсти, что позволило сделать более сильный укус[15][16][17].
Традиционно считалось, что черепахи являются выжившими парарептилиями на основании их анапсидной структуры черепа, которая считалась примитивной чертой. Обоснование этой классификации оспаривалось, некоторые утверждали, что черепахи являются диапсидами, которые развили анапсидные черепа, чтобы улучшить свою броню. Более поздние морфологические филогенетические исследования с учётом этого поместили черепах прочно в состав Diapsida. Все молекулярные исследования убедительно подтвердили принадлежность черепах к диапсидам, чаще всего как сестринскую группу по отношению к существующим архозаврам[18][19][20][21][22].
Пермские рептилии[править | править код]
С окончанием карбона амниоты стали доминирующими. В то время как примитивные наземные рептилиоморфы всё ещё существовали, синапсидные амниоты представили мегафауну (гигантских животных) в форме пеликозавров, таких как эдафозавр и плотоядный диметродон. В середине пермского периода климат стал более сухим, что привело к изменению фауны: на смену пеликозаврам пришли терапсиды[23].
Парарептилии, чьи массивные черепа не имели заглазничных отверстий, сохранялись и процветали на протяжении всего пермского периода. Парарептилии из числа парейазавров достигли гигантских размеров в поздней перми и в конце концов исчезли в конце периода (возможно, выжили черепахи[23]).
В начале этого периода современные рептилии развились и разделились на две основные линии: архозавроморфы (предки черепах, крокодилов и динозавров) и лепидозавроморфы (предшественники современных ящериц и туатары). Обе группы оставались ящерицеподобными, относительно небольшими и незаметными в течение пермского периода[23].
Мезозойские рептилии[править | править код]
В конце пермского периода произошло крупнейшее известное массовое вымирание (см. Пермско-триасовое вымирание)[24]. Большая часть более ранней парарептильной и синапсидной мегафауны исчезла, заменившись похожими на современных рептилиями, особенно архозавроморфами. Они характеризовались длинными задними ногами и прямохождением, ранние формы чем-то напоминали длинноногих крокодилов. Архозавры стали доминирующей группой в триасовый период, хотя потребовалось 30 миллионов лет, прежде чем их разнообразие стало таким же большим, как у животных, живших в пермском периоде[24]. Архозавры превратились в хорошо известных нам динозавров и птерозавров, а также предков крокодилов. Поскольку рептилии, сначала рауизухии, а затем динозавры, доминировали в мезозойскую эру, этот интервал широко известен как «эпоха рептилий». Динозавры были как гигантскими, так и мелкими (в том числе более мелких теропод с перьями). В меловой период они дали начало первым настоящим птицам[25].
Сестринской группой Archosauromorpha является Lepidosauromorpha, в которую входят ящерицы и туатары, а также их ископаемые родственники. Lepidosauromorpha содержал по крайней мере одну основную группу мезозойских морских рептилий: мозазавров, живших в меловой период. Более спорным является филогенетическое размещение других основных групп ископаемых морских рептилий — ихтиоптеригов (включая ихтиозавров) и завроптеригов, которые эволюционировали в раннем триасе. Разные авторы связывали эти группы либо с лепидозавроморфами, либо с архозавроморфами. Итиоптериги также считались диапсидами, которые не принадлежали к наименее инклюзивной кладе, содержащей лепидозавроморфов и архозавроморфов[24][26][27].
Кайнозойские (современные) пресмыкающиеся[править | править код]
В конце мелового периода произошла гибель мегафауны рептилий мезозойской эры (см. Мел-палеогеновое вымирание, знаменитое исчезновением динозавров). Из крупных морских рептилий остались только морские черепахи; а из неморских крупных рептилий только полуводные крокодилы и в целом похожие на них хористодеры пережили вымирание, а последние представители последних, похожие на ящериц Lazarussuchus, вымерли в миоцене. Из огромного множества динозавров, господствовавших в мезозое, выжили только мелкие клювовидные птицы. Эта драматическая модель вымирания в конце мезозоя перешла в кайнозой. Млекопитающие и птицы заполнили пустые ниши, оставленные рептилиями, и, в то время как диверсификация рептилий замедлилась, диверсификация птиц и млекопитающих приняла экспоненциальный оборот. Тем не менее, рептилии по-прежнему были важными организмами фауны[28][29][30].
После исчезновения большинства линий архозавров и морских рептилий к концу мелового периода диверсификация рептилий продолжалась на протяжении всего кайнозоя. Много видов чешуйчатых вымерло во время Мел-палеогенового вымирания, восстановившись только через десять миллионов лет после него. Сегодня чешуйчатые составляют большинство живых рептилий (> 95 %). Известно примерно 11 950 современных видов традиционных рептилий, а птиц ещё около 10 000, что вчетверо превышает количество млекопитающих, представленных примерно 5700 живыми видами[31][32][33][34].
Классификация[править | править код]
В XIII веке категория рептилий была признана в Европе как состоящая из множества яйцекладущих существ, включая «змей, различных фантастических чудовищ, ящериц, различных амфибий и червей», как записано Винсентом де Бове в его «Зеркале»[35]. В XVIII веке рептилии с самого начала классификации были отнесены к амфибиям. Карл Линней, работавший в бедной видами Швеции, где обыкновенная гадюка и ужей уж часто охотятся в воде, включил всех рептилий и земноводных в класс «III — Амфибии» в его Systema Naturæ[36]. Термины «рептилия» и «амфибия» были в значительной степени взаимозаменяемы, рептилия (от латинского repere , «ползать») предпочиталась французами. Иосиф Флавий Николаус Лауренти был первым, кто официально использовал термин Reptilia для обозначения рептилий и амфибий, в основном сходного с тем, что у Линнея. Сегодня эти две группы всё ещё изучаются одной наукой герпетологией[37][38].
Только в начале XIX века стало ясно, что рептилии и земноводные на самом деле совершенно разные животные, и Пьер Андре Латрейль возвёл для последних класс Baracia (1825 г.), разделив четвероногих на четыре известных класса: рептилий, земноводных, птиц и млекопитающих. Британский анатом Томас Генри Хаксли сделал определение Латрейля популярным и вместе с Ричардом Оуэном расширил Reptilia, включив в него различных ископаемых допотопных монстров”, включая динозавров и млекопитающих (синапсид) Dicynodon. Это была не единственная возможная схема классификации: в лекциях Хантера, прочитанных в Королевском колледже хирургов в 1863 году, Хаксли сгруппировал позвоночных в млекопитающих, завроидов и ихтиоидов (последние включали рыб и земноводных). Впоследствии он предложил названия Sauropsida и Ichthyopsida для двух последних групп. В 1866 году Геккель продемонстрировал, что позвоночных можно разделить на основе их репродуктивных стратегий, и что рептилий, птиц и млекопитающих объединяет амниотическое яйцо[39].
Термины Sauropsida («лица ящериц») и Theropsida («лица зверей») были снова использованы в 1916 г. Э. С. Гудричем для различения ящериц, птиц и их родственников, с одной стороны (Sauropsida), и млекопитающих и их вымерших родственников (Theropsida) с другой. Гудрич поддержал это разделение по характеру сердца и сосудов в каждой группе и другим особенностям, таким как строение переднего мозга. По словам Гудрича, обе линии произошли от более ранней основной группы, Protosauria («первые ящерицы»), в которую он включил некоторых животных, которые сегодня считаются рептилиеподобными амфибиями, а также ранних рептилий[40].
В 1956 году Дэвид Мередит Сирс Уотсон заметил, что первые две группы разошлись очень рано в истории рептилий, поэтому он разделил Protosauria Гудрича между ними. Он также переосмыслил Sauropsida и Theropsida, исключив соответственно птиц и млекопитающих. Таким образом, его Sauropsida включала Procolophonia, Eosuchia, Millerosauria, Chelonia (черепахи), Squamata (ящерицы и змеи), Rhynchocephalia, Crocodilia, «текодонтов» (парафилетические базальные Archosauria), нептичьих динозавров, птерозавров, ихтиозавров и завроптериги[41].
В конце XIX века был предложен ряд переопределений Рептилий. Признаки, перечисленные Лидеккером в 1896 году, например, включают единственный затылочный мыщелок, челюстной сустав, образованный квадратной и суставной костями, и некоторые характеристики позвонков. Животные, выделяемые этими формулировками, амниоты, отличные от млекопитающих и птиц, до сих пор считаются рептилиями[42].
Разделение синапсид/завропсид дополнило другой подход, который разделил рептилий на четыре подкласса. Эта классификация была инициирована Генри Фэйрфилдом Осборном и развита и стала популярной благодаря классической книге «Палеонтология позвоночных» Ромера. Этими четырьмя подклассами были[43][44]:
- Anapsida — котилозавры и Chelonia (черепахи и их родственники)
- Synapsida — пеликозавры и терапсиды («звероподобные рептилии»)
- Эвриапсиды — проторозавры (маленькие, ранние ящероподобные рептилии) и морские завроптериги и ихтиозавры, последних в работе Осборна назвали Парапсидами.
- Diapsida — большинство рептилий, включая ящериц, змей, крокодилов, динозавров и птерозавров.
Расположение Euryapsida было неопределённым. Иногда считалось, что ихтиозавры возникли независимо от других эвриапсидов, и им дали более старое название Парапсида. Позже Parapsida была по большей части исключена как группа (ихтиозавры были классифицированы как incertae sedis вместе с Euryapsida). Однако четыре (или три, если Euryapsida сливаются с Diapsida) подкласса оставались более или менее универсальными для неспециалистов на протяжении всего XX века. Недавние исследователи в значительной степени отказались от него[45].
К началу XXI века палеонтологи позвоночных начали принимать филогенетическую таксономию, в которой все группы определяются таким образом, чтобы быть монофилетическими; то есть группы, в которые входят все потомки определённого предка. Рептилии, как исторически определено, являются парафилетическими, поскольку они исключают как птиц, так и млекопитающих. Они соответственно произошли от динозавров и от ранних терапсидов, которых традиционно называли рептилиями. Птицы более тесно связаны с крокодилами, чем последние с остальными современными рептилиями[46]. Несмотря на ранние предложения о замене парафилетических рептилий монофилетическими завропсидами, включающими птиц, этот термин так и не получил широкого распространения, а если и применялся, то не применялся последовательно[47].
Колин Тадж писал:
Млекопитающие являются кладой, и поэтому кладисты рады признать традиционный таксон млекопитающих; и птицы тоже представляют собой кладу, повсеместно приписываемую формальному таксону Aves. Mammalia и Aves, по сути, являются субкладами в большой кладе Amniota. Но традиционный класс Reptilia не является кладой. Это всего лишь часть клады Amniota: часть, оставшаяся после отделения Mammalia и Aves. Он не может быть определён синапоморфно, как это правильно. Вместо этого он определяется комбинацией свойств, которые у него есть, и признаков, которых ему не хватает: рептилии — это амниоты, у которых нет меха или перьев. В лучшем случае, предполагают кладисты, мы могли бы сказать, что традиционные рептилии — это «амниоты, не относящиеся к птицам и млекопитающим»[42].
Mammals are a clade, and therefore the cladists are happy to acknowledge the traditional taxon Mammalia; and birds, too, are a clade, universally ascribed to the formal taxon Aves. Mammalia and Aves are, in fact, subclades within the grand clade of the Amniota. But the traditional class Reptilia is not a clade. It is just a section of the clade Amniota: the section that is left after the Mammalia and Aves have been hived off. It cannot be defined by synapomorphies, as is the proper way. Instead, it is defined by a combination of the features it has and the features it lacks: reptiles are the amniotes that lack fur or feathers. At best, the cladists suggest, we could say that the traditional Reptilia are ‘non-avian, non-mammalian amniotes’.
Когда использовался термин Sauropsida, она имел почти то же определение, что и Reptilia. В 1988 году Жак Готье предложил кладистическое определение Reptilia как монофилетической группы, включающей черепах, ящериц и змей, крокодилов и птиц, их общего предка и всех его потомков. Хотя определение Готье было близко к современному консенсусу, тем не менее, оно стало считаться неверным, потому что фактическая связь черепах с другими рептилиями в то время ещё не была хорошо изучена. С тех пор основные изменения включали переназначение синапсид как нерептилий и классификацию черепах как диапсид[46].
Множество других определений было предложено другими учёными в годы после статьи Готье. Первое такое новое определение, было опубликовано Модесто и Андерсоном в 2004 году. Они определили Reptilia как всех амниот, более близких к Lacerta agilis и Crocodylus niloticus, чем к Homo sapiens. Это основанное на стебле определение эквивалентно более распространённому определению Sauropsida, которое Модесто и Андерсон синонимизировали с Reptilia, поскольку последнее более известно и чаще используется. Однако, в отличие от большинства предыдущих определений Reptilia, определение Модесто и Андерсона включает птиц, поскольку они входят в кладу, включающую как ящериц, так и крокодилов[46].
Дочерние таксоны[править | править код]
Класс Пресмыкающиеся (Reptilia)[48][49][50]
- † Подкласс Парарептилии (Parareptilia) — возможно, базальные диапсиды[51]
- † Отряд Мезозавры (Mesosauria)
- † Отряд Millerosauria
- † Отряд Проколофономорфы (Procolophonomorpha)
- † ? Отряд Болозавры (Bolosauria)[51] — возможно, в отряде Procolophonomorpha[52]
- Подкласс Диапсиды (Diapsida)
- Отряд † Ареосцелидии (Araeoscelidia)
- Клада Неодиапсиды (Neodiapsida)
- Отряд † Талаттозавры (Thalattosauria)
- Клада † Drepanosauromorpha
- Клада † Ichthyosauromorpha — положение спорно
- Надотряд † Ихтиоптеригии (Ichthyopterygia)
- Отряд † Ихтиозавры (Ichthyosauria)
- Надотряд † Ихтиоптеригии (Ichthyopterygia)
- Клада Заврии (Sauria)
- incertae sedis
- Отряд † Хористодеры (Choristodera) — возможно, базальные архозавроморфы[53]
- Надотряд † Завроптеригии (Sauropterygia) — возможно, парафилетическая группа к Testudinata[53]
- Отряд † Плакодонты (Placodontia)
- Отряд † Nothosauroidea
- Отряд † Плезиозавры (Plesiosauria)
- Инфракласс Лепидозавроморфы (Lepidosauromorpha)
- Надотряд Лепидозавры (Lepidosauria)
- Отряд Клювоголовые (Rhynchocephalia)
- Отряд Чешуйчатые (Squamata)
- Надотряд Лепидозавры (Lepidosauria)
- Инфракласс Архозавроморфы (Archosauromorpha s. l.)[54] [ ≈ Archelosauria[55][56]]
- Клада Тестудинаты (Testudinata)
- Отряд Черепахи (Testudines)
- Клада Архозавроморфы (Archosauromorpha s. s.)[57]
- Клада Crocopoda[58]
- Клада † Allokotosauria[59][60]
- Отряд † Ринхозавры (Rhynchosauria)
- Клада Архозавриформы (Archosauriformes)
- Отряд † Фитозавры (Phytosauria) и более базальные таксоны[61][62]
- Клада Архозавры (Archosauria)
- Клада Псевдозухии (Pseudosuchia)
- Отряд † Этозавры (Aetosauria) и прочие базальные таксоны
- Надотряд Крокодиломорфы (Crocodylomorpha)
- Отряд Крокодилы (Crocodilia)
- Клада Авеметатарзалии (Avemetatarsalia)[63]
- Клада † Aphanosauria
- Клада Орнитодиры (Ornithodira)
- Отряд † Птерозавры (Pterosauria)
- Надотряд Динозавры (Dinosauria)
- Отряд Ящеротазовые (Saurischia) — в кладистике включает птиц (Aves)
- Отряд † Птицетазовые (Ornithischia)
- Клада Псевдозухии (Pseudosuchia)
- Клада Crocopoda[58]
- Клада Тестудинаты (Testudinata)
- incertae sedis
Филогенетика[править | править код]
Согласно определению Modesto & Anderson, 2004, пресмыкающиеся, или рептилии (Reptilia), являются наиболее инклюзивной кладой, включающей Lacerta agilis Linnaeus, 1758 и Crocodylus niloticus Laurenti, 1768, но не Homo sapiens Linnaeus, 1758. Данное определение эквивалентно определению завропсид (Sauropsida), данному Готье в 1994 году, согласно которому к этой кладе относятся все пресмыкающиеся и другие амниоты, ближе родственные им, чем млекопитающим (Mammalia)[4][64].
Кладограмма согласно Lee, 2013[53]:
Кладограмма, основанная на Ford & Benson, 2020[51]:
Строение[править | править код]
Пресмыкающиеся, наряду с птицами и млекопитающими относятся к высшим позвоночным животным — амниотам, и в целом их строение типично для данной группы, однако, также имеются черты более простых по строению амфибий.
Покров[править | править код]



Наружный кожный покров современных пресмыкающихся в результате утолщения и ороговения образует чешуйки или щитки. У ящериц роговые чешуйки перекрывают друг друга, напоминая черепицу. У черепах сросшиеся щитки формируют сплошной прочный панцирь. Смена рогового покрова происходит путём полной или частичной линьки, которая у многих видов происходит несколько раз в год.
Наряду с этим, древние рептилии могли также быть покрыты своеобразными перьями (являющимися по строению специализированной формой чешуи) или похожими на пух нитевидными образованиями (прогрессивные архозавры).
Плотная и сухая кожа содержит пахучие железы. Слизистые железы отсутствуют — кожа рептилий сухая, характерный блеск ей придают гладкие чешуи.
В наружной части внутреннего слоя кожи часто находятся специальные клетки — хроматофоры. В этих клетках секретируются пигменты: меланины и каротиноиды. Также в хроматофорах встречается способный отражать свет гуанин. Благодаря хроматофорам, некоторые пресмыкающиеся способны изменять окраску своего тела за сравнительно короткое время. Хамелеоны — наиболее известные представители с подобным свойством.
Скелет[править | править код]
В осевом скелете пресмыкающихся разделение на отделы более заметно, чем у земноводных. Хорошо различимы пять отделов позвоночника: шейный (лат. pars cervicalis), туловищный (пояснично-грудной, pars thoracolumbalis), поясничный, крестцовый (pars sacralis) и хвостовой (pars caudalis).
Типичным для пресмыкающихся является следующее строение осевого скелета. Общее количество позвонков различно у разных видов (50—80, у змей возрастает до 140—435). Из позвонков шейного отдела (от 7 до 10) два передних (атлант и эпистрофей) образуют сустав, позволяющий голове не только двигаться в вертикальной плоскости относительно первого шейного позвонка, но и поворачиваться. В туловищном отделе от 16 до 25 позвонков, каждый с парой рёбер. Первые несколько пар рёбер прикрепляются к грудине, образуя грудную клетку (отсутствует у змей). В крестцовом отделе всего два позвонка, к широким поперечным отросткам которых причленяется таз. Хвостовой отдел составляют несколько десятков (15—40) постепенно уменьшающихся в размерах позвонков. Последние хвостовые позвонки представляют собой небольшие палочковидные косточки.
В некоторых группах рептилий осевой скелет имеет отличия. У змей позвоночник отчётливо делится лишь на туловищный и хвостовой отделы, грудина отсутствует. У черепах позвонки туловищного отдела срастаются со спинным щитом панциря, вследствие чего неподвижны.
Череп пресмыкающихся значительно более окостеневший, чем у земноводных. Лишь в обонятельной капсуле и слуховой области содержится небольшое количество хряща. Осевой и висцеральный отделы черепа эмбрионально формируются отдельно, но у взрослых особей срастаются в единое образование. В состав черепа входят как хрящевые (замещающие, или первичные), так и многочисленные кожные (покровные, или вторичные) кости.
Пояс передних конечностей сходен с поясом земноводных, отличаясь лишь более сильным развитием окостенения. Пара передних конечностей рептилий состоит из плеча, предплечья и кисти. Пара задних конечностей — из бедра, голени и стопы. На фалангах конечностей расположены когти.
Мышечная система[править | править код]
Мышечная система пресмыкающихся представлена жевательной, шейной мускулатурой, мускулатурой брюшного пресса, а также мускулатурой сгибателей и разгибателей. Присутствуют характерные для амниот межрёберные мышцы, играющие важную роль при акте дыхания. Подкожная мускулатура позволяет изменять положение роговых чешуй.
Энергоснабжение произвольной мускулатуры современных пресмыкающихся происходит главным образом за счёт анаэробного разложения глюкозы — порядка 50—75 % мышечных волокон в их мышцах приспособлены к работе в анаэробных условиях (без участия кислорода). Такой метаболизм позволяет им перемещаться на короткие расстояния столь же быстро, как и теплокровным животным, и обеспечивает даже большую силу сокращения мышц, причём интенсивность реакции у многих групп слабо зависит от температуры тела. Однако уже через 1—2 минуты интенсивной работы мускулатуры в анаэробном режиме в ней накапливается молочная кислота, что приводит к наступлению своеобразной мышечной усталости и изменению pH крови. В результате уже после нескольких быстрых пробежек рептилия становится практически неспособна к активным действиям на время от нескольких минут до нескольких часов, требующееся для распада накопившейся в мышцах молочной кислоты[65]. Более крупные животные способны дольше терпеть накопление в мышцах молочной кислоты и таким образом фактически являются более выносливыми. Крокодилы способны выдерживать падение pH крови до 6,42 и ниже[66].
Некоторые современные рептилии, в частности, вараны и кожистая черепаха, тем не менее, являются исключением из этого правила: в ходе активности их уровень метаболизма может достигать более половины от такового у теплокровных животных сопоставимой массы, и они в значительной мере полагаются на аэробный метаболизм, почти не испытывая проблем с накоплением молочной кислоты[67].
Среди вымерших древних рептилий известны высокоподвижные формы (такие как архозавры: наземные крокодиломорфы, динозавры и птерозавры, а также лепидозавры: мозазавры, ихтиозавры и завроптеригии), строение тела и предполагаемый образ жизни которых говорят о способности поддерживать продолжительную мышечную активность без утомления, что соответствует более интенсивному, чем у современных пресмыкающихся, обмену веществ, и работе мышц преимущественно по аэробному механизму, требующему интенсивного снабжения кислородом. Это в свою очередь должно было сопровождаться совершенно другой физиологией организма (полное разделение желудочков сердца, повышение эффективности дыхания за счёт отделения воздухоносных путей от ротовой полости, появление эффективных механизмов терморегуляции и так далее). Данные изменения отчасти прослеживаются на палеонтологическом материале в линии рептилий, ведущей к птицам. В настоящее время надёжно показано существование теплокровности как минимум у некоторых групп динозавров и предков современных крокодилов, а также плезиозавров, мозазавров и ихтиозавров[64][65][68].
Нервная система[править | править код]

Как и у большинства хордовых животных, центральная нервная система пресмыкающихся представлена головным (из 5 отделов) и спинным мозгом.
Головной мозг расположен внутри черепа. Ряд важных особенностей отличает головной мозг пресмыкающихся от головного мозга земноводных. Нередко говорят[кто?] о так называемом завропсидном типе головного мозга, присущем также и птицам, в отличие от ихтиопсидного типа у рыб и земноводных.
Выделяют пять отделов головного мозга пресмыкающихся.
- Передний мозг состоит из двух больших полушарий, от которых отходят обонятельные доли. Поверхность больших полушарий абсолютно гладкая. В мозговом своде полушарий различают первичный свод — архипаллиум, занимающий большую часть крыши полушарий, и зачатки неопаллиума. Дно переднего мозга в основном состоит из полосатых тел.
- Промежуточный мозг расположен между передним и средним мозгом. В верхней его части расположен эпифиз, а на нижней стороне — гипофиз. У большинства ящериц и гаттерии (а также у многих вымерших форм) рядом с эпифизом развивается теменной глаз, а у крокодилов потеряны оба этих органа. Дно промежуточного мозга занято зрительными нервами и их перекрестом (хиазмой).
- Средний мозг представлен двумя крупными передними холмами — зрительными долями, а также небольшими задними холмами. Зрительная кора более развита, чем у амфибий.
- Мозжечок прикрывает переднюю часть продолговатого мозга. Он более крупный по сравнению с мозжечком амфибий.
- Продолговатый мозг образует изгиб в вертикальной плоскости, который характерен для всех амниот.
От головного мозга отходят 12 пар черепных нервов. В спинном мозге разделение на белое и серое вещество более отчётливо, чем у амфибий. От спинного мозга отходят сегментальные спинномозговые нервы, образуя типичное плечевое и тазовое сплетение. Отчётливо выражена вегетативная нервная система (симпатическая и парасимпатическая) в виде цепи парных нервных ганглиев.
Органы чувств[править | править код]

Пресмыкающиеся имеют шесть основных органов чувств, из которых зрение имеет решающее значение.
- Орган зрения — глаза, устроены сложнее, чем у амфибий: в склере присутствует кольцо из тонких костных пластинок; от задней стенки глазного яблока отходит вырост — гребешок, вдающийся в стекловидное тело; в ресничном теле развита поперечно-полосатая мускулатура, которая позволяет не только перемещать хрусталик, но и изменять его форму, таким образом осуществляя наводку на резкость в процессе аккомодации. Органы зрения имеют приспособления к работе в воздушной среде. Слёзные железы предохраняют глаза от высыхания. Наружные веки и мигательная перепонка выполняют защитную функцию. У змей и некоторых ящериц веки срастаются, формируя прозрачную оболочку. Сетчатка глаза может содержать как палочки, так и колбочки. У ночных видов колбочки отсутствуют. У большинства дневных видов диапазон цветного зрения смещён в жёлто-оранжевую часть спектра. Форма зрачка чаще всего круглая или в виде вертикальной щели (как у кошки), горизонтальный щелевидный зрачок, весьма распространённый у земноводных, среди рептилий встречается очень редко, например у плетевидных змей. Теменной глаз, сохранившийся у некоторых видов, важен для ориентирования в пространстве и синхронизации суточных ритмов организма с циклом смены дня и ночи, хотя многое в его функциях ещё неясно[69][70].
- Орган обоняния представлен внутренними ноздрями — хоанами и вомероназальным органом. По сравнению со строением земноводных, хоаны расположены ближе к глотке, что даёт возможность свободно дышать в то время, как пища находится во рту. Обоняние развито лучше, чем у земноводных, позволяя многим ящерицам находить пищу, находящуюся под поверхностью песка на глубине до 6—8 см.
- Орган вкуса — вкусовые луковицы, расположенные в основном в глотке.
- Орган тепловой чувствительности находится на лицевой ямке между глазом и носом с каждой стороны головы. Особенно развит у змей. У ямкоголовых змей термолокаторы позволяют определять даже направление источника теплового излучения.
- Орган слуха близок к органу слуха лягушек, он содержит внутреннее и среднее ухо, снабжённое барабанной перепонкой, слуховой косточкой — стременем и евстахиевой трубой. Роль слуха в жизни пресмыкающихся сравнительно невелика, особенно слаб слух у змей, не имеющих барабанной перепонки и воспринимающих колебания, распространяющиеся по земле или в воде. Пресмыкающиеся воспринимают звуки в диапазоне 20—6000 Гц, хотя большинство хорошо слышит лишь в диапазоне 60—200 Гц (у крокодилов 100—3000 Гц).
- Осязание выражено отчётливо, особенно у черепах, которые могут чувствовать даже лёгкое прикосновение к панцирю.
Дыхательная система[править | править код]
Для пресмыкающихся характерно дыхание всасывающего типа путём расширения и сужения грудной клетки при помощи межрёберной и брюшной мускулатуры. Попавший через гортань воздух поступает в трахею — длинную дыхательную трубку, которая на конце делится на бронхи, ведущие в лёгкие. Как и у земноводных, лёгкие пресмыкающихся имеют мешкообразное строение, хотя их внутренняя структура намного сложнее. Внутренние стенки лёгочных мешков имеют складчатое ячеистое строение, что значительно увеличивает дыхательную поверхность. Строение лёгких значительно отличается у разных видов и может представлять слабодифференциированные органы с малым количеством ячей, у примитивных видов, таких как гаттерии, среднеразвитые органы с ярко выраженной внутренней структурой, как у большинства чешуйчатых, так и высокоразвитые лёгкие, имеющие почти губчатое строение (как у птиц и млекопитающих), у таких рептилий, как вараны, тегу, крокодилы, многие черепахи и крупные змеи. Водные черепахи имеют более проницаемую кожу, а некоторые виды даже изменили свою клоаку, чтобы увеличить площадь газообмена. Даже с этими адаптациями дыхание никогда полностью не осуществляется без лёгких[71]. Смена воздуха в лёгких у каждой основной группы рептилий осуществляется по-разному. У чешуйчатых лёгкие вентилируются почти исключительно осевой мускулатурой. Это та же самая мускулатура, которая используется при передвижении. Из-за этого ограничения большинство чешуек вынуждены задерживать дыхание во время интенсивных пробежек. Некоторые, однако, нашли способ обойти это. Вараниды и некоторые другие виды ящериц используют буккальное сцеживание как дополнение к их обычному «аксиальному дыханию». Это позволяет животным полностью наполнять свои лёгкие во время бега и, таким образом, длительное время оставаться аэробно активными. Известно, что у ящериц тегу имеется протодиафрагма, отделяющая лёгочную полость от висцеральной. Хотя на самом деле он не способен к движению, он позволяет сильнее раздувать лёгкие, снимая вес внутренних органов с них[72].
Поскольку тело покрыто чешуйками, кожное дыхание у пресмыкающихся отсутствует (исключение составляют мягкотелые черепахи и морские змеи), и лёгкие являются единственным дыхательным органом.
У видов, живущих в воде, могут появляться специализированные адаптации, для того чтобы обходиться дольше без атмосферного воздуха, например у морских змей это слизистая выстилка рта, а у черепах это и полость рта и специальные анальные мешки, способные усваивать кислород из воды.
Крокодилы на самом деле имеют мышечную диафрагму, аналогичную диафрагме млекопитающих. Разница в том, что мышцы крокодиловой диафрагмы тянут лобковую кость (часть таза, подвижную у крокодилов) назад, что опускает печень, тем самым освобождая пространство для расширения лёгких. Этот тип диафрагмальной установки называют «печёночным поршнем». Дыхательные пути образуют ряд двойных трубчатых камер в каждом лёгком. При вдохе и выдохе воздух проходит через дыхательные пути в одном направлении, создавая таким образом однонаправленный поток воздуха через лёгкие. Аналогичная система обнаружена у птиц, варанов и игуан[73][74].
У большинства рептилий нет вторичного неба, а это означает, что они должны задерживать дыхание при глотании. Крокодилы развили костлявое вторичное нёбо, которое позволяет им продолжать дышать, оставаясь под водой (и защищать свой мозг от повреждений, связанных с борьбой с добычей). Сцинки (семейство Scincidae) также в той или иной степени развили костное вторичное небо. Змеи использовали другой подход и вместо этого расширили свои трахеи. Их расширение трахеи торчит изо рта в виде трубки, и позволяет этим животным проглатывать крупную добычу, не страдая от удушья[75].
Система кровообращения[править | править код]
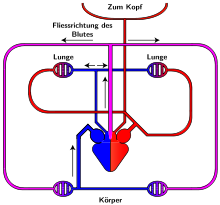
Система кровообращения пресмыкающихся
Как и амфибии, большинство пресмыкающихся обладают трёхкамерным сердцем, состоящим из желудочка и двух предсердий. Желудочек разделён неполной перегородкой на две половины: верхнюю и нижнюю, и может быть условно разделён на три отдела. Однако трёхкамерное сердце рептилий развилось независимо от такового у земноводных, и различается в плане строения (и иногда — функциональности) у разных современных групп[76][77].
Все лепидозавры и черепахи имеют трёхкамерное сердце, состоящее из двух предсердий, одного желудочка с переменными перегородками и двух аорт, ведущих в большой круг кровообращения. Степень смешения оксигенированной и деоксигенированной крови в трёхкамерном сердце различна в зависимости от вида и физиологического состояния. При различных условиях деоксигенированная кровь может быть отправлена обратно в организм, а насыщенная кислородом кровь может быть отправлена обратно в лёгкие. Было высказано предположение, что это изменение кровотока обеспечивает более эффективную терморегуляцию и более длительное время погружения для водных видов, но не было доказано, что оно является преимуществом и для наземных видов[78].
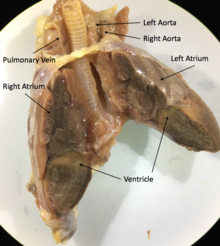
Будучи архозаврами, крокодилы обладают четырёхкамерным сердцем[64]. Перегородка полностью разделяет желудочек на две половины: правую — венозную и левую — артериальную, таким образом обосабливая полноценное четырёхкамерное сердце, как у млекопитающих и птиц. Лишь после образования полных перегородок у крокодилов образуется отверстие Панница в ходе эмбриогенеза, допускающее контролируемое смешивание артериальной и венозной крови, что связано со вторичным понижением уровня обмена веществ у этой группы животных[64]. Крокодилы имеют анатомически четырёхкамерное сердце, похожее на птичье, но также имеют две аорты[79].
Например, сердце игуаны, как и большинство чешуйчатых сердец, состоит из трёх камер с двумя аортами и одним желудочком, сердечными непроизвольными мышцами. Основными структурами сердца являются венозный синус, водитель ритма, левое предсердие, правое предсердие, атриовентрикулярный клапан, венозная полость, артериальная полость, лёгочная полость, мышечный гребень, вентрикулярный гребень, лёгочные вены и парные дуги аорты[80].
Некоторые виды чешуйчатых (например, питоны и вараны) имеют трёхкамерное сердце, которое при сокращении функционально становится четырёхкамерным. Это стало возможным благодаря мышечному гребню, который разделяет желудочек во время диастолы желудочков и полностью разделяет его во время систолы желудочков . Из-за этого гребня некоторые из этих чешуек способны создавать перепады давления в желудочках, эквивалентные тем, которые наблюдаются в сердцах млекопитающих и птиц[81].
Полного разделения на два независимых круга кровообращения у пресмыкающихся не происходит, поскольку венозная и артериальная кровь смешиваются в спинной аорте.
Подобно рыбам и амфибиям, подавляющее большинство современных пресмыкающихся — холоднокровные животные. Несмотря на это, многие пресмыкающиеся могут регулировать температуру, перемещаясь из тени на солнце и обратно или меняя цвет, становясь темнее, чтобы согреться, или светлее, чтобы остыть. Сравнительно недавно у ящериц тегу была обнаружена факультативная эндотермия — возможность повышать температуру тела во время брачного периода за счёт эндотермических химических реакций[82]. Вараны в разгар пищевой активности также могут повышать температуру тела за счёт внутренних процессов[83]. Кожистая черепаха, в свою очередь, имеет изолирующую жировую прослойку и за счёт интенсивной мышечной активности может быть примерно на 18 градусов теплее окружающей её воды[84].
Метаболизм[править | править код]
Современные нептичьи рептилии холоднокровны, так что они имеют ограниченные физиологические средства поддержания постоянной температуры тела и часто полагаются на внешние источники тепла. Из-за менее стабильной внутренней температуры, чем у птиц и млекопитающих, биохимия рептилий требует ферментов, способных сохранять эффективность в более широком диапазоне температур, чем в случае теплокровных .животные. Оптимальный диапазон температуры тела зависит от вида, но обычно ниже, чем у теплокровных животных; для многих ящериц она находится в диапазоне 24-35 °C (75-95 °F), в то время как у экстремально адаптированных к жаре видов, таких как американская пустынная игуана Dipsosaurus dorsalis, могут быть оптимальные физиологические температуры для млекопитающих, в диапазоне от 35° до 40 °C (от 95° до 104 °F). В то время как оптимальная температура часто наблюдается, когда животное активно, низкий основной обмен приводит к быстрому падению температуры тела, когда животное неактивно[85][86].
Как и у всех животных, мышечная деятельность рептилий производит тепло. У крупных рептилий, таких как кожистые черепахи, низкое отношение поверхности к объёму позволяет этому метаболически вырабатываемому теплу держать животных теплее, чем их окружающая среда, даже если у них нет теплокровного метаболизма. Эта форма гомеотермии называется гигантотермией; было высказано предположение, что она была обычным явлением у крупных динозавров и других вымерших крупных рептилий[87].
Преимущество низкого метаболизма в состоянии покоя заключается в том, что для поддержания функций организма требуется гораздо меньше пищи. Используя колебания температуры окружающей среды или оставаясь холодными, когда им не нужно двигаться, рептилии могут экономить значительное количество энергии по сравнению с эндотермическими животными того же размера. Крокодилу требуется от одной десятой до одной пятой части пищи, необходимой льву того же веса, и он может прожить без еды полгода. Более низкие потребности в пище и адаптивный метаболизм позволяют рептилиям доминировать в животной жизни в регионах, где доступность пищи слишком мала для поддержания крупных млекопитающих и птиц[88][89].
Принято считать, что рептилии не могут производить устойчиво высокий уровень энергии, необходимый для погони на большие расстояния или полёта. Более высокая энергетическая ёмкость могла быть ответственна за эволюцию теплокровности у птиц и млекопитающих. Однако исследование корреляции между активной ёмкостью и термофизиологией показывает слабую связь. Большинство современных рептилий — плотоядные животные со стратегией кормления «сиди и жди»; неясно, являются ли рептилии хладнокровными из-за их экологии. Энергетические исследования некоторых рептилий показали, что их активные способности равны или превосходят возможности теплокровных животных аналогичного размера[90][91].
Пищеварительная система[править | править код]
Вследствие разнообразия доступной пищи[прояснить] пищеварительный тракт у пресмыкающихся гораздо более дифференцирован, чем у земноводных животных.
Пищеварительная система начинается ротовым отверстием, ограниченным челюстями с обычно коническими, одинаковыми, прочно прирастающими зубами (гомодонтная система, хотя у некоторых вымерших форм была гетеродонтная). У рептилий, за исключением крокодилов и большинства черепах, есть гибкие неподвижные губы, плотно закрывающие рот и зубы в состоянии покоя. Зубы пресмыкающихся не предназначены для жевания — они служат для захвата и удержания пищи, реже — для её разделения или отрывания удобных по размеру кусков, у видов, питающихся моллюсками и насекомыми, зубы приспособлены для дробления жёстких покровов этих животных. Пища практически всегда проглатывается целиком, как животная, так и растительная. Язык свободный, впереди мускулистый, подвижный, к концу утончается и раздваивается. У некоторых видов язык клейкий, и служит для ловли добычи. Язык служит для проглатывания пищи, но виды с тонким и длинным языком проталкивают пищу в глотку толчками головы (ящерицы) или движением челюстей (змеи) Ротовая полость отграничена от глотки развивающимся вторичным костным нёбом у ящериц и черепах, полностью развитым костным небом у крокодилов, но у змей и варанов вторичное костное небо отсутствует. Многоклеточные слюнные железы содержат пищеварительные ферменты. Глотка переходит в узкий, но очень растяжимый пищевод, далее в мускулистый желудок и кишечник. Желудок имеет толстые мышечные стенки. На границе между тонкой и толстой кишкой находится слепая кишка, которой нет у амфибий. Большая печень рептилий имеет жёлчный пузырь. Поджелудочная железа в виде длинного плотного тела лежит в петле двенадцатиперстной кишки. Кишечник заканчивается клоакой.
Выделительная система[править | править код]

Фекалии ящерицы с кристаллами мочевой кислоты
Выделительная система пресмыкающихся представлена почками, мочеточниками и мочевым пузырём.
Почки пресмыкающихся существенно отличаются от почек рыб и амфибий, которым приходится решать задачу избавления от постоянного избытка воды в организме. Вместо туловищных почек амфибий (мезонефрос), почки рептилий (метанефрос) располагаются в тазовой области с брюшной стороны клоаки и по её бокам. Почки соединяются с клоакой через мочеточники.
Тонкостенный стебельчатый мочевой пузырь соединяется с клоакой тонкой шейкой на её брюшной стороне. У некоторых пресмыкающихся мочевой пузырь недоразвит (крокодилы, змеи, некоторые ящерицы).
У наземных рептилий в качестве конечного продукта азотистого обмена выступает слабо растворимая в воде мочевая кислота, выделяемая вместе с калом через клоаку, что позволяет более эффективно сохранять влагу.
Половая система[править | править код]
Пресмыкающиеся — раздельнополые животные, двуполое размножение.
Половая система самцов состоит из пары семенников, которые расположены по бокам поясничного отдела позвоночника. От каждого семенника отходит семенной канал, который впадает в вольфов канал. С появлением тазовой почки у пресмыкающихся вольфов канал у самцов выступает лишь как семяпровод и полностью отсутствует у самок. Вольфов канал открывается в клоаку, образуя семенной пузырёк.
Половая система самок представлена яичниками, которые подвешены на брыжейке к спинной стороне полости тела по бокам позвоночника. Яйцеводы (мюллеровы каналы) также подвешены на брыжейке. В переднюю часть полости тела яйцеводы открываются щелевидными отверстиями — воронками. Нижний конец яйцеводов открывается в нижний отдел клоаки на её спинной стороне.
Образ жизни[править | править код]
Развитие[править | править код]
Оплодотворение внутреннее. Развитие зародыша происходит в яйце с кожистой или известковой оболочкой, наряду с этим встречается яйцеживорождение и (реже) истинное живорождение. У пресмыкающихся прямое постэмбриональное развитие. Для многих представителей характерна забота о потомстве, в частности самки крокодилов переносят потомство от места кладки к водоёмам в ротовой полости, хотя в некоторых случаях могут съесть детёныша.
Питание[править | править код]
Большинство пресмыкающихся относится к плотоядным животным (включая насекомоядность и поедание падали, истинное хищничество среди современных рептилий свойственно только крокодилам, ряду ящериц и большинству змей). Для некоторых (например, агамы, игуаны) характерно смешанное питание. Иногда встречаются и почти исключительно травоядные пресмыкающиеся (сухопутные черепахи).
Передвижение[править | править код]

Протерозух, ранний архозавроморф с конечностями, расположенными по бокам от туловища, как у современных рептилий, и приспособленными к «пресмыканию».

Примитивные группы древних амниот (в том числе первые рептилии) изначально передвигались на четырёх широко расставленных в стороны конечностях, которые при локомоции раздвигались медиалатерально и не были способны постоянно поддерживать туловище над землёй, отсюда — «пресмыкание», волочение живота по земле, давшее русскоязычное название всей этой группе, что соответствовало характерному для них низкому уровню интенсивности обмена веществ и сравнительно малоподвижному образу жизни, связанному с чередованием коротких вспышек активности и длительного отдыха лёжа на животе. Современные пресмыкающиеся в основном сохраняют подобный, базальный в эволюционном плане, характер локомоции, некоторое исключение составляют только крокодилы, у которых конечности при беге могут занимать положение почти под туловищем. Причём у некоторых вымерших групп современного отряда крокодилов, которые вели исключительно сухопутный образ жизни, такое положение конечностей было габитуальным, то есть поддерживалось большую часть времени — в Австралии такие наземные крокодилы (например, Квинкана) обитали ещё 40 тыс. лет назад, то есть одновременно с человеком современного вида. У варанов конечности полувыпрямлены, что связано с активным перемещением, и при беге туловище находится высоко над землёй. Змеи и безногие ящерицы так же является исключением, так как отсутствие конечностей и совершенно иной способ локомоции (ползание), чем у рептилий с конечностями, обеспечивают быстрое перемещение большинству змей по любой поверхности.
Большинство рептилий неплохо плавают за счёт «рыбообразного» изгибания тела в поперечной плоскости. У некоторых групп приспособление к водной среде достигло более высокой степени, причём в процессе эволюции у них вырабатывались различные типы локомоции. Так, ихтиозавры, мозазавры и, в меньшей степени, крокодилы сохранили характер плавания за счёт поперечных изгибов туловища. С другой стороны, у морских черепах и вымерших плезиозавров выработался новый механизм движения в воде, за счёт взмахов конечностей, преобразовавшихся в ласты. Многие эти группы практически полностью утратили способность перемещаться на суше. Несколько родов пресмыкающихся способны к планирующему полёту — например летучие драконы (агамы рода Draco) и украшенные древесные змеи. Активно летающие рептилии известны только из ископаемых остатков (см. Птерозавры).
В древности развитие некоторых продвинутых групп рептилий шло в иных направлениях — их конечности стали перемещаться непосредственно под туловище, занимая более габитуально вертикальное (выпрямленное) положение, что соответствовало более подвижному образу жизни, связанному с быстрым бегом и активным преследованием добычи.

У предков архозавров лобковая и седалищная кости начали удлиняться, передние ноги — укорачиваться, а тяжёлый длинный хвост, практически лишившийся подвижности из-за срастания тазовых костей, превратился в балансир, уравновешивающий массу вытянутого туловища. Задние конечности при ходьбе стали перемещаться в парасагиттальной плоскости, голеностопный сустав получил возможность пружинить при ходьбе, что способствовало более экономному расходу сил, а опора переместилась на средний (третий) палец. Это позволило многим группам текодонтов перейти к факультативной, или даже габитуальной, бипедальности, подобно современным бегающим птицам, специализировавшись на беге на двух ногах.
Данные изменения должны были сопровождаться значительным повышением интенсивности обмена веществ, поскольку постоянное поддержание тела над землёй и длительная локомоторная активность требовали намного больших энергозатрат по сравнению с «пресмыканием». Однако проследить подобные изменения на палеонтологическом материале сложно. Достоверно известно, однако, что потомки данной группы рептилий — птицы — имеют интенсивный обмен веществ и эндотермную систему терморегуляции (теплокровность).
Ранние динозавры, как ящеротазовые, так и птицетазовые — были активными хищниками, как минимум часть времени передвигавшимися на задних конечностях, что способствовало их быстрому прогрессу, особенно после вымирания большинства синапсидов на рубеже пермского и триасового периода. В юрском периоде почти все более-менее крупные наземные животные (свыше метра в длину) были динозаврами.
Постепенно часть динозавров переходит на растительную пищу, занимая освободившиеся экологические ниши. Большинство растительноядных видов вновь возвращается к передвижению на четырёх конечностях, эволюция же хищников продолжает идти по «двуногому» пути (например, у тираннозаврид и абелизаврид передние конечности оказались практически редуцированны, а у манирапторов — напротив, развились в специализированную хватательную форму). В течение 140 млн лет, вплоть до конца мелового периода, основными наземными хищными позвоночными были двуногие динозавры, некоторые формы которых по большинству признаков неотличимы от птиц.
Как видно, исторически для рептилий было свойственно главным образом именно активное передвижение на двух или четырёх выпрямленных конечностях, именно такие бегающие формы занимали господствующее положение в данном классе (парафилетической группе) животных на протяжении большей части его истории. Двуногие хищные формы изредка возникали у птиц — например фороракосы. В настоящее время подобные формы среди рептилий не возникают, поскольку экологические ниши, связанные с активным преследованием добычи, ныне заняты млекопитающими, и организм современных рептилий требует значительной перестройки для их освоения. Некоторое исключение составляет насекомоядная плащеносная ящерица, и некоторые другие агамовые, способные к бегу на двух ногах (при сохранении расположения конечностей по бокам от туловища). Такая особенность позволяет им обходить ограничение Карриера (невозможность бежать на четырёх ногах и дышать одновременно) и активно преследовать добычу или спасаться от хищников. Ящерица шлемоносный василиск во время бипедальной локомоции способна также бегать по поверхности воды. Тегу и вараны также научились обходить ограничение Карриера, но за счёт модификации дыхательной системы, а не подъёма на задние конечности.
Издаваемые звуки[править | править код]
Большинство рептилий не имеет настоящего голосового аппарата и производят лишь самые примитивные звуки вроде шипения или свиста. Крокодилы рычат, издают лающие и рокочущие звуки, детёныши квакают. Черепахи в некоторых случаях (испуг, брачный сезон, боль) могут издавать писк или стон. Представители гекконовых ящериц издают самые разные звуки — писк, визг, щебет, щелчки, крики[92].
Хозяйственное значение[править | править код]
Значение пресмыкающихся для человека сравнительно невелико. Кожа крокодилов, крупных змей и ящериц применяется в кожевенной промышленности для изготовления чемоданов, ремней, обуви и т. п., однако эти предметы носят эксклюзивный характер, являясь предметом роскоши. Мясо и яйца многих черепах употребляются в пищу. Также могут употребляться в пищу некоторые ящерицы и змеи. Змеиный яд находит применение в медицине. Многие змеи полезны истреблением грызунов, а ящерицы — насекомых. Некоторые виды пресмыкающихся держат в качестве домашних животных. Аллигаторы, кайманы и некоторые крокодилы иногда разводятся на специальных фермах, для получения мяса, яиц и кожи.
Большую опасность для людей представляют ядовитые змеи, особенно в тропических странах. Многие крокодилы опасны для человека, наносят значительный ущерб животноводству. Пресноводные черепахи вредят рыбному промыслу. Крупные вараны, питоны и удавы так же могут быть опасны, будучи способными травмировать или даже убить человека.
Модельные объекты[править | править код]
В 2011 году был расшифрован геном пресмыкающегося — ящерицы анолис каролинский[93]. Таким образом, это пресмыкающееся вошло в круг модельных объектов генетики.
См. также[править | править код]
- Пресмыкающиеся, описанные в 2010 году
Примечания[править | править код]
- ↑ Laurin M., Reisz R. R. A reevaluation of early amniote phylogeny (англ.) // Zoological Journal of the Linnean Society (англ.) (рус. : journal. — 1995. — Vol. 113, no. 2. — P. 165—223. — doi:10.1111/j.1096-3642.1995.tb00932.x. Архивировано 8 июня 2019 года.
- ↑ Gauthier J. A., Kluge A. G., Rowe T. The early evolution of the Amniota // The Phylogeny and Classification of the Tetrapods (англ.) / Benton M. J.. — Oxford: Oxford University Press, 1988. — Vol. 1. — P. 103—155. — ISBN 978-0-19-857705-8.
- ↑ Modesto S. P. Observations of the structure of the Early Permian reptile Stereosternum tumidum Cope (англ.) // Palaeontologia Africana : journal. — 1999. — Vol. 35. — P. 7—19.
- ↑ 1 2 Modesto S. P., Anderson J. S. The phylogenetic definition of Reptilia (англ.) // Systematic Biology : journal. — 2004. — Vol. 53, no. 5. — P. 815—821. — doi:10.1080/10635150490503026. — PMID 15545258.
- ↑ Species Numbers (as of December 2022) (англ.). Reptile Database. Дата обращения: 6 января 2023. Архивировано 6 января 2023 года.
- ↑ Высшие таксоны животных: данные о числе видов для России и всего мира Архивировано 21 февраля 2012 года..
- ↑ См. также Список пресмыкающихся России.
- ↑ A reevaluation of early amniote phylogeny // Zoological Journal of the Linnean Society.
- ↑ Science News Online (5/22/99): Out of the Swamps. web.archive.org (4 июня 2011). Дата обращения: 10 сентября 2022.
- ↑ Earliest Evidence For Reptiles / Scientific Frontline®. web.archive.org (16 июля 2011). Дата обращения: 10 сентября 2022.
- ↑ The illustrated encyclopedia of dinosaurs and prehistoric animals. — London: Marshall Pub, 1999. — С. 62. — 312 pages с. — ISBN 1-84028-152-9, 978-1-84028-152-1.
- ↑ 1 2 3 Neil Brocklehurst. The First Age of Reptiles? Comparing Reptile and Synapsid Diversity, and the Influence of Lagerstätten, During the Carboniferous and Early Permian // Frontiers in Ecology and Evolution. — 2021. — Т. 9. — ISSN 2296-701X. — doi:10.3389/fevo.2021.669765/full.
- ↑ Links between global taxonomic diversity, ecological diversity and the expansion of vertebrates on land // Biology letters.
- ↑ Rainforest collapse triggered Carboniferous tetrapod diversification in Euramerica. pubs.geoscienceworld.org. Дата обращения: 10 сентября 2022.
- ↑ Graciela Piñeiro, Jorge Ferigolo, Alejandro Ramos, Michel Laurin. Cranial morphology of the Early Permian mesosaurid Mesosaurus tenuidens and the evolution of the lower temporal fenestration reassessed (англ.) // Comptes Rendus Palevol. — 2012-07-01. — Vol. 11, iss. 5. — P. 379–391. — ISSN 1631-0683. — doi:10.1016/j.crpv.2012.02.001.
- ↑ Juan C. Cisneros, Ross Damiani, Cesar Schultz, Atila da Rosa, Cibele Schwanke, Leopoldo W. Neto, and Pedro L. P. Aurélio. A procolophonoid reptile with temporal fenestration from the Middle Triassic of Brazil // Proc Biol Sci.
- ↑ M. Tuinen, E. Hadly. Error in Estimation of Rate and Time Inferred from the Early Amniote Fossil Record and Avian Molecular Clocks // Journal of Molecular Evolution. — 2004. — doi:10.1007/s00239-004-2624-9.
- ↑ M. J. Benton. Vertebrate palaeontology. — 3rd ed. — Malden, MA: Blackwell Science, 2005. — xi, 455 pages с. — ISBN 0-632-05637-1, 978-0-632-05637-8.
- ↑ O. Rieppel, M. Debraga. Turtles as diapsid reptiles // Nature. — 1996. — doi:10.1046/j.1463-6409.2000.00039.x.
- ↑ Rafael Zardoya and Axel Meyer. Complete mitochondrial genome suggests diapsid affinities of turtles // Proc Natl Acad Sci U S A.
- ↑ Sister Group Relationship of Turtles to the Bird-Crocodilian Clade Revealed by Nuclear DNA–Coded Proteins. academic.oup.com. Дата обращения: 11 сентября 2022.
- ↑ Y. Katsu, E. Braun, L. Guillette Jr, T. Iguchi. From Reptilian Phylogenomics to Reptilian Genomes: Analyses of c-Jun and DJ-1 Proto-Oncogenes // Cytogenetic and Genome Research. — 2009. — doi:10.1159/000297715.
- ↑ 1 2 3 Edwin H. Colbert. Colbert’s evolution of the vertebrates : a history of the backboned animals through time. — 5th ed. — New York: Wiley, 2001. — xvi, 560 pages с. — ISBN 0-471-38461-5, 978-0-471-38461-8.
- ↑ 1 2 3 Sarda Sahney and Michael J. Benton. Recovery from the most profound mass extinction of all time // Proc Biol Sci.
- ↑ Morphological Clocks in Paleontology, and a Mid-Cretaceous Origin of Crown Aves. academic.oup.com. Дата обращения: 11 сентября 2022.
- ↑ Abstracts of Papers // Journal of Vertebrate Paleontology. — 1997-09-04. — Т. 17, вып. sup003. — С. 1–93. — ISSN 0272-4634. — doi:10.1080/02724634.1997.10011028.
- ↑ Ryosuke Motani, Nachio Minoura, Tatsuro Ando. Ichthyosaurian relationships illuminated by new primitive skeletons from Japan (англ.) // Nature. — 1998-05. — Vol. 393, iss. 6682. — P. 255–257. — ISSN 1476-4687. — doi:10.1038/30473.
- ↑ Susan E. Evans, Jozef Klembara. [0171:ACRRDF2.0.CO;2 A choristoderan reptile (Reptilia: Diapsida) from the Lower Miocene of Northwest Bohemia (Czech Republic)] // Journal of Vertebrate Paleontology. — 2005-03-11. — Т. 25, вып. 1. — С. 171–184. — ISSN 0272-4634. — doi:10.1671/0272-4634(2005)025[0171:ACRRDF]2.0.CO;2.
- ↑ Dennis M. Hansen, C. Josh Donlan, Christine J. Griffiths, Karl J. Campbell. Ecological history and latent conservation potential: large and giant tortoises as a model for taxon substitutions (англ.) // Ecography. — 2010-06-15. — P. no–no. — doi:10.1111/j.1600-0587.2010.06305.x.
- ↑ Alberto Cione, Eduardo Tonni, Leopoldo Soilbenzon. The Broken Zig-Zag: Late Cenozoic large mammal and tortoise extinction in South America // Revista del Museo Argentino de Ciencias Naturales nueva serie. — 2014-06-16. — Т. 5, вып. 1. — С. 1–19. — ISSN 1853-0400.
- ↑ Nicholas R. Longrich, Bhart-Anjan S. Bhullar and Jacques A. Gauthiera. Mass extinction of lizards and snakes at the Cretaceous–Paleogene boundary // Proc Natl Acad Sci U S A.
- ↑ Reptile Database News. www.reptile-database.org. Дата обращения: 11 сентября 2022.
- ↑ Tod W. Reeder, Ted M. Townsend, Daniel G. Mulcahy, Brice P. Noonan, Perry L. Wood, Jr., Jack W. Sites, Jr., and John J. Wiens. Integrated Analyses Resolve Conflicts over Squamate Reptile Phylogeny and Reveal Unexpected Placements for Fossil Taxa // PLoS One.
- ↑ Table 1: Numbers of threatened species by major groups of organisms (1996–2012). web.archive.org (4 февраля 2013). Дата обращения: 11 сентября 2022.
- ↑ Mary Franklin-Brown. Reading the world : encyclopedic writing in the scholastic age. — Chicago: The University of Chicago Press, 2012. — 1 online resource с. — ISBN 978-0-226-26070-9, 0-226-26070-4.
- ↑ Carl von Linné, Lars Salvius. Caroli Linnaei…Systema naturae per regna tria naturae :secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. — Holmiae: Impensis Direct. Laurentii Salvii, 1758. — 881 с.
- ↑ Amphibia (1878). aleph0.clarku.edu. Дата обращения: 10 сентября 2022.
- ↑ Josephi Nicolai Laurenti specimen medicum exhibens synopsin reptilium emendatam cum experimentis circa venena et antidota reptilium Austriacorum.
- ↑ The Structure and Classification of the Mammalia (1864). aleph0.clarku.edu. Дата обращения: 10 сентября 2022.
- ↑ Edwin Stephen Goodrich. On the classification of the reptilia // Proceedings of the Royal Society of London. Series B, Containing Papers of a Biological Character. — 1916-07-01. — Т. 89, вып. 615. — С. 261–276. — doi:10.1098/rspb.1916.0012.
- ↑ David Meredith Seares Watson. On Millerosaurus and the early history of the sauropsid reptiles // Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. — 1957-03-28. — Т. 240, вып. 673. — С. 325–398. — doi:10.1098/rstb.1957.0003.
- ↑ 1 2 Colin Tudge. The variety of life : a survey and a celebration of all the creatures that have ever lived. — Oxford: Oxford University Press, (2002 [printing]). — xiv, 684 pages с. — ISBN 0-19-860426-2, 978-0-19-860426-6.
- ↑ Romer, A.S. Vertebrate Paleontology. — University of Chicago Press, 1933.
- ↑ Osborn, H.F. The Reptilian subclasses Diapsida and Synapsida and Early History of Diaptosauria. — Memoirs of the American Museum of Natural History, 1903. — С. 451–507.
- ↑ L. A. Tsuji, J. Müller. Assembling the history of the Parareptilia: phylogeny, diversification, and a new definition of the clade (англ.) // Fossil Record. — 2009-02-01. — Т. 12, вып. 1. — С. 71–81. — ISSN 2193-0066. — doi:10.1002/mmng.200800011.
- ↑ 1 2 3 Keynyn Brysse. From weird wonders to stem lineages: the second reclassification of the Burgess Shale fauna // Studies in History and Philosophy of Biological and Biomedical Sciences. — 2008-09. — Т. 39, вып. 3. — С. 298–313. — ISSN 1369-8486. — doi:10.1016/j.shpsc.2008.06.004.
- ↑ Assembling the history of the Parareptilia: phylogeny, diversification, and a new definition of the clade. academic.oup.com. Дата обращения: 10 сентября 2022.
- ↑ Benton M. J.ruen, Spencer P. S. Fossil Reptiles of Great Britain (англ.). — Chapman & Hall, 1995. — P. 8. — 386 p. — ISBN 978-94-010-4231-4. — ISBN 978-94-011-0519-4.
- ↑ Benton M. J. Vertebrate palaeontologyruen (англ.). — 4th ed. — Wiley-Blackwell, 2015. — P. 437. — 437—441 p. — ISBN 978-1-118-40755-4. — ISBN 978-1-118-40684-7.
- ↑ Sues H.-D.ruen. The Rise of Reptiles: 320 Million Years of Evolution (англ.). — Baltimore: Johns Hopkins University Press, 2019. — P. ix—xi. — 385 p. — ISBN 9781421428680. — ISBN 1421428687.
- ↑ 1 2 3 Ford D. P., Benson R. B. J. The phylogeny of early amniotes and the affinities of Parareptilia and Varanopidae (англ.) // Nature Ecology & Evolution : journal. — 2020. — Vol. 4, iss. 1. — P. 57–65. — ISSN 2397-334X. — doi:10.1038/s41559-019-1047-3. Архивировано 18 сентября 2020 года.
- ↑ MacDougall M. J., Reisz R. R. The first record of a nyctiphruretid parareptile from the Early Permian of North America, with a discussion of parareptilian temporal fenestration (англ.) // Zoological Journal of the Linnean Society. — 2014. — Vol. 172, iss. 3. — P. 616—630. — ISSN 1096-3642. — doi:10.1111/zoj.12180. Архивировано 2 сентября 2021 года.
- ↑ 1 2 3 Lee M. S. Y. Turtle origins: insights from phylogenetic retrofitting and molecular scaffolds (англ.) // Journal of Evolutionary Biology. — 2013. — Vol. 26, iss. 12. — P. 2729–2738. — ISSN 1420-9101. — doi:10.1111/jeb.12268. Архивировано 8 января 2022 года.
- ↑ Gauthier J. A., Kluge A. G., Rowe T. B. Amniote Phylogeny and the Importance of Fossils (англ.) // Cladisticsruen : journal. — 1988. — Vol. 4, iss. 2. — P. 105—209. — ISSN 1096-0031. — doi:10.1111/j.1096-0031.1988.tb00514.x. Архивировано 1 сентября 2021 года.
- ↑ Crawford N. G., Parham J. F., Sellas A. B., et al. A phylogenomic analysis of turtles (англ.) // Molecular Phylogenetics and Evolution : journal. — 2015. — Vol. 83. — P. 250—257. — ISSN 1055-7903. — doi:10.1016/j.ympev.2014.10.021.
- ↑ Joyce W. G., Anquetin J., Cadena E.-A., et al. A nomenclature for fossil and living turtles using phylogenetically defined clade names (англ.) // Swiss Journal of Palaeontology. — 2021. — Vol. 140, iss. 1. — P. 5. — ISSN 1664-2384. — doi:10.1186/s13358-020-00211-x. Архивировано 3 апреля 2021 года.
- ↑ de Queiroz, Cantino & Gauthier, 2020, Archosauromorpha F. von Huene 1946 [J. A. Gauthier], converted clade name, pp. 1179—1182.
- ↑ Spiekman S. N. F., Fraser N. C., Scheyer T. M. A new phylogenetic hypothesis of Tanystropheidae (Diapsida, Archosauromorpha) and other “protorosaurs”, and its implications for the early evolution of stem archosaurs (англ.) // PeerJruen : journal. — 2021. — Vol. 9. — P. e11143. — ISSN 2167-8359. — doi:10.7717/peerj.11143. — PMID 33986981. Архивировано 14 августа 2021 года.
- ↑ Nesbitt S. J., Flynn J. J., Pritchard A. C., Parrish M. J., Ranivoharimanana L., Wyss A. R. Postcranial osteology of Azendohsaurus madagaskarensis (?Middle to Upper Triassic, Isalo Group, Madagascar) and its systematic position among stem archosaur reptiles (англ.) // Bulletin of the American Museum of Natural Historyruen : journal. — 2015. — Vol. 398. — P. 1—126. — ISSN 0003-0090. — doi:10.5531/sd.sp.15. Архивировано 13 июля 2021 года.
- ↑ Sengupta S., Ezcurra M. D., Bandyopadhyay S. A new horned and long-necked herbivorous stem-archosaur from the Middle Triassic of India (англ.) // Scientific Reportsruen : journal. — 2017. — Vol. 7, iss. 1. — P. 8366. — ISSN 2045-2322. — doi:10.1038/s41598-017-08658-8. Архивировано 2 сентября 2021 года.
- ↑ Nesbitt S. J. The early evolution of archosaurs: relationships and the origin of major clades (англ.) // Bulletin of the American Museum of Natural History : journal. — 2011. — Vol. 352. — P. 1—292. — ISSN 0003-0090. — doi:10.1206/352.1. Архивировано 20 августа 2021 года.
- ↑ Marsh A. D., Smith M. E., Parker W. G., Irmis R. B., Kligman B. T. Skeletal anatomy of Acaenasuchus geoffreyi Long and Murry, 1995 (Archosauria: Pseudosuchia) and its implications for the origin of the aetosaurian carapace (англ.) // Journal of Vertebrate Paleontology. — 2020. — Vol. 40, iss. 4. — P. e1794885. — ISSN 0272-4634. — doi:10.1080/02724634.2020.1794885. Архивировано 1 сентября 2021 года.
- ↑ Nesbitt S. J., Butler R. J., Ezcurra M. D., et al. The earliest bird-line archosaurs and the assembly of the dinosaur body plan (англ.) // Nature : journal. — 2017. — Vol. 544, iss. 7651. — P. 484—487. — ISSN 1476-4687. — doi:10.1038/nature22037. Архивировано 28 мая 2019 года.
- ↑ 1 2 3 4 Seymour, R. S., Bennett-Stamper, C. L., Johnston, S. D., Carrier, D. R. and Grigg, G. C. Evidence for endothermic ancestors of crocodiles at the stem of archosaur evolution : [англ.] // Physiological and Biochemical Zoology. — 2004. — Vol. 77, no. 6. — P. 1051—1067. — doi:10.1086%2F422766 10.1086/422766. — PMID 15674775. Источник. Дата обращения: 14 ноября 2017. Архивировано 19 октября 2017 года..
- ↑ 1 2 Кэрролл Р. . Палеонтология и эволюция позвоночных: В 3 т. Т. 2. — М.: Мир, 1993. — 283 с. — ISBN 5-03-001819-0.
- ↑ G. J. W. Webb, D. F. Bradford, R. S. Seymour, A. F. Bennett. Mass-Dependence of Anaerobic Metabolism and Acid-Base Disturbance During Activity in the Salt-Water Crocodile, Crocodylus Porosus (англ.) // The Journal of Experimental Biology. — The Company of Biologists (англ.) (рус., 1985-09-01. — Vol. 118, iss. 1. — P. 161—171. — ISSN 0022-0949 1477-9145, 0022-0949. Архивировано 10 декабря 2018 года.
- ↑ Seymour, Roger S.; Smith, Sarah L; White, Craig R.; Henderson, Donald M.; Schwarz-Wings, Daniela. Blood flow to long bones indicates activity metabolism in mammals, reptiles and dinosaurs : [англ.] // Proceedings of the Royal Society B. — 2012. — Vol. 279, no. 1728. — P. 451—456. — doi:10.1098/rspb.2011.0968. — PMID 21733896. — PMC 3234558.
- ↑ Giant Sea Reptiles Were Warm-Blooded? (12 июня 2010). Архивировано 9 декабря 2018 года. Дата обращения: 31 мая 2018.
- ↑ Labra A., Voje K. L., Seligmann H., Hansen T. F. Evolution of the third eye: a phylogenetic comparative study of parietal-eye size as an ecophysiological adaptation in Liolaemus lizards (англ.) // Biological Journal of the Linnean Society (англ.) (рус. : journal. — Oxford University Press, 2010. — Vol. 101, no. 4. — P. 870—883. — doi:10.1111/j.1095-8312.2010.01541.x.
- ↑ Tosini G. The pineal complex of reptiles: physiological and behavioral roles (англ.) // Ethology Ecology & Evolution : journal. — 1997. — Vol. 9, no. 4. — P. 313—333. — doi:10.1080/08927014.1997.9522875.
- ↑ Ronald I. (Ronald Isaac) Orenstein. Turtles, tortoises and terrapins : survivors in armor. — Buffalo, N.Y. : Firefly Books, 2001. — 330 с. — ISBN 978-1-55209-605-5.
- ↑ W. Klein, A. Abe, D. Andrade, S. Perry. Structure of the posthepatic septum and its influence on visceral topology in the tegu lizard, Tupinambis merianae (Teiidae: Reptilia) // Journal of morphology. — 2003. — doi:10.1002/JMOR.10136.
- ↑ New insight into the evolution of the vertebrate respiratory system and the discovery of unidirectional airflow in iguana lungs // Proc Natl Acad Sci U S A.
- ↑ C. Farmer, K. Sanders. Unidirectional Airflow in the Lungs of Alligators // Science. — 2010. — doi:10.1126/science.1180219.
- ↑ Gross anatomy of the snake.
- ↑ А. Ромер, Т. Парсонс. Анатомия позвоночных. Том 1 Альфред Ромер, Томас Парсонс. log-in.ru. Дата обращения: 31 мая 2018. Архивировано 12 июня 2018 года.
- ↑ Анатомия позвоночных. Том 2 — Биологический кружок – Юные Исследователи Природы. uipmgu.ru. Дата обращения: 31 мая 2018. Архивировано 11 июля 2018 года.
- ↑ J. Hicks. The physiological and evolutionary significance of cardiovascular shunting patterns in reptiles. // News in physiological sciences : an international journal of physiology produced jointly by the International Union of Physiological Sciences and the American Physiological Society. — 2002. — doi:10.1152/NIPS.01397.2002.
- ↑ Michael Axelsson, Craig E. Franklin. From Anatomy to Angioscopy: 164 Years of Crocodilian Cardiovascular Research, Recent Advances, and Speculations (англ.) // Comparative Biochemistry and Physiology Part A: Physiology. — 1997-09-01. — Vol. 118, iss. 1. — P. 51–62. — ISSN 0300-9629. — doi:10.1016/S0300-9629(96)00255-1.
- ↑ “Reptilian cardiovascular anatomy and physiology: evaluation and monitoring (Proceedings).
- ↑ Ventricular haemodynamics in Python molurus: separation of pulmonary and systemic pressures. journals.biologists.com. Дата обращения: 11 сентября 2022.
- ↑ Glenn J. Tattersall, Cleo A. C. Leite, Colin E. Sanders, Viviana Cadena, Denis V. Andrade. Seasonal reproductive endothermy in tegu lizards (англ.) // Science Advances. — 2016-01-01. — Vol. 2, iss. 1. — P. e1500951. — ISSN 2375-2548. — doi:10.1126/sciadv.1500951. Архивировано 2 июля 2017 года.
- ↑ Jorge Cubo, Meike Köhler, Vivian de Buffrénil. [https://onlinelibrary.wiley.com/doi/abs/10.1111/let.12203 Bone histology of Iberosuchus macrodon
(Sebecosuchia, Crocodylomorpha)] (англ.) // Lethaia. — 2017-03-27. — Vol. 50, iss. 4. — P. 495—503. — ISSN 0024-1164. — doi:10.1111/let.12203. Архивировано 5 июня 2020 года. - ↑ Frair W, Ackman RG, Mrosovsky N. Body Temperatures of Dermochelys coriacea: Warm Turtle from Cold Water : [англ.] // Science. — 1972. — Vol. 177, no. 4051. — P. 791—793. — doi:10.1126/science.177.4051.791. — PMID 17840128.
- ↑ Raymond B. Huey; Albert F. Bennett. [http://faculty.washington.edu/hueyrb/pdfs/Coadaptation_Evolution.pdf Phylogenetic Studies of Coadaptation: Preferred Temperatures Versus
Optimal Performance Temperatures of Lizards] // Evolution. - ↑ RAYMOND B. HUEY. Temperature, Physiology, and the Ecology of Reptiles // Department of Zoology, University of Washington, Seattle, U.S.A..
- ↑ The complete dinosaur. — Bloomington: Indiana University Press, 1997. — С. 491–504. — xi, 752 pages с. — ISBN 0-253-33349-0, 978-0-253-33349-0, 0-253-21313-4, 978-0-253-21313-6.
- ↑ Neil A. Campbell. Essential biology. — 2nd ed. — San Francisco: Pearson/Benjamin Cummings, 2004. — С. 396. — xxxi, 464, [44] pages с. — ISBN 0-8053-7495-7, 978-0-8053-7495-7, 0-321-20462-X, 978-0-321-20462-2, 0-8053-7473-6, 978-0-8053-7473-5, 0-8053-7478-7, 978-0-8053-7478-0, 0-321-22537-6, 978-0-321-22537-5.
- ↑ S. T. Garnett. Metabolism and survival of fasting Estuarine crocodiles (англ.) // Journal of Zoology. — 1986-04. — Vol. 208, iss. 4. — P. 493–502. — ISSN 1469-7998 0952-8369, 1469-7998. — doi:10.1111/j.1469-7998.1986.tb01518.x.
- ↑ Pat Willmer. Environmental physiology of animals. — Malden, MA: Blackwell Science, 2000. — С. 444. — x, 644 pages, 10 unnumbered pages of plates с. — ISBN 0-632-03517-X, 978-0-632-03517-5.
- ↑ James W. Hicks, Colleen G. Farmer. Gas exchange potential in reptilian lungs: implications for the dinosaur–avian connection (англ.) // Respiration Physiology. — 1999-09-15. — Vol. 117, iss. 2. — P. 73–83. — ISSN 0034-5687. — doi:10.1016/S0034-5687(99)00060-2.
- ↑ Морозов В. П. Занимательная биоакустика. Изд. 2-е, доп., перераб. — М.: Знание, 1987. — 208 с. + 32 с. вкл. — С. 63-65
- ↑ Рудый, Юлия Рудый Впервые расшифрован геном пресмыкающегося. membrana.ru (5 сентября 2011). Дата обращения: 6 сентября 2011. Архивировано 13 ноября 2011 года.
Литература[править | править код]
- Наумов Н. П., Карташев Н. Н. Часть 2. Пресмыкающиеся, птицы, млекопитающие // Зоология позвоночных. — М.: Высшая школа, 1979. — С. 272.
- Карташев Н. Н., Соколов В. Е., Шилов И. А. Практикум по зоологии позвоночных. — М.: Высшая школа, 1981.
- Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989.
- Жизнь животных. Энциклопедия в шести томах. Том 5. (Земноводные. Пресмыкающиеся). Общая редакция члена-корреспондента АН СССР профессора Л. А. Зенкевича. — Москва: Просвещение, 1969. — 488 стр.
- Phylonyms: A Companion to the PhyloCode (англ.) / de Queiroz K.ruen, Cantino P. D., Gauthier J. A., eds. — Boca Raton: Taylor & Francis Group, CRC Press, 2020. — 1352 p. — ISBN 978-1-138-33293-5.
Ссылки[править | править код]
- Пресмыкающиеся // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
- Классификация современных пресмыкающихся Архивная копия от 13 февраля 2012 на Wayback Machine (англ.)
Пресмыкающиеся, или рептилии – класс наземных позвоночных животных типа хордовые, размножение и развитие
которых более не зависят (как у земноводных) от водоема. Относятся к амниотам. Рептилии достигли наибольшего расцвета в мезозое.
На настоящий момент класс насчитывает около 10 тысяч видов.

Рептилии произошли от древних панцирноголовых земноводных – стегоцефалов, которые приспособились к защите тела от высыхания и
смогли освоить отдаленные от водоемов пространства.

Независимость от водоема досталась пресмыкающимся с трудом, для этого их организм приобрел ряд прогрессивных черт – ароморфозов:
- Зародышевые оболочки
- Внутреннее оплодотворение
- Сухая кожа
- Ячеистые легкие и грудная клетка
- Неполная перегородка в сердце
- Кора больших полушарий
- Вторичная почка
Как сказано выше, пресмыкающиеся относятся к амниотам. Это значит, что развитие их зародышей происходит в особом зародышевом пузыре
с амниотической жидкостью – амнионе. Поразительно! Им более не нужен водоем, этот водоем всегда с собой, прямо в организме самки вокруг зародыша.
К зародышевым оболочкам относится наружная, серозная оболочка (сероза), внутренняя амниотическая оболочка и аллантоис.
Аллантоис – зародышевый орган, с помощью которого осуществляется дыхание.
Эти оболочки позволили пресмыкающимся расселиться от водоемов вглубь материков, занять огромные территории.
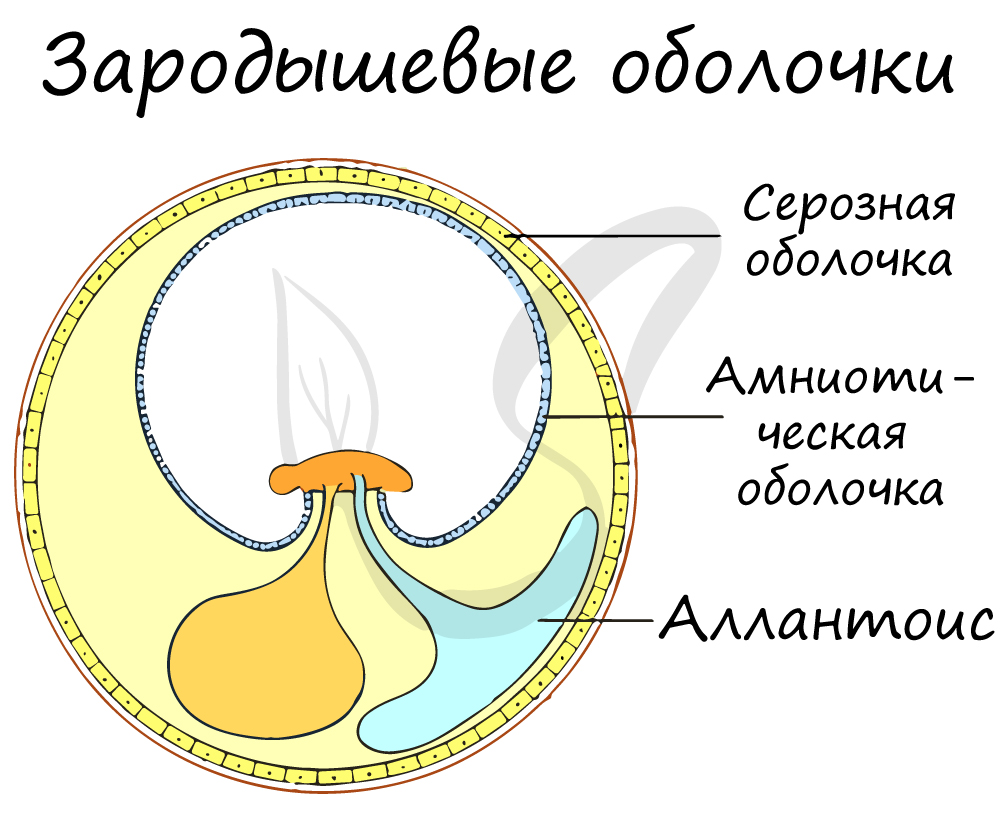
У рыб и земноводных оплодотворение было наружным, малоэффективным, с небольшой вероятностью встречи половых клеток. При
внутреннем оплодотворении, которое появляется у рептилий, семенная жидкость самца со сперматозоидами вводится непосредственно в
половые пути самки, что значительно увеличивает вероятность встречи мужских и женских гамет.

Кожа пресмыкающихся сухая, практически лишена желез (которых так много у земноводных). Верхние слои эпителия ороговевают,
на поверхности кожи формируются роговые чешуйки. Эта защита необходима от пересыхания, помогает эффективнее удерживать воду
в организме.

У пресмыкающихся происходит усложнение строения дыхательной системы: появляются ячеистые легкие, обеспечивающие более
эффективный газообмен. Кожное дыхание (доминировавшее у амфибий) у пресмыкающихся сведено к минимуму или отсутствует вовсе.
Дыхательные пути у рептилий дифференцируются на гортань, трахею и бронхи.
Дыхание реберного типа – в нем участвуют появившиеся вместе с ребрами межреберные мышцы. Ребра образуют
новую структуру скелета – грудную клетку.
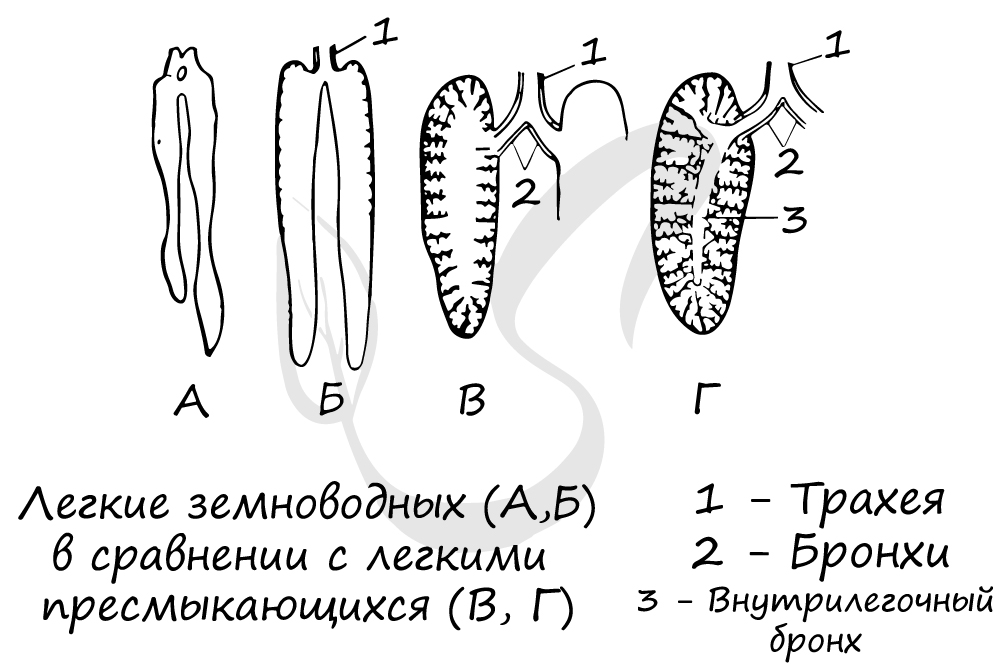
В сердце у пресмыкающихся развивается неполная межжелудочковая перегородка, способствующая более эффективному разделению
артериальной и венозной крови. Это, в свою очередь, повышает эффективность обмена веществ (син. – метаболизм).
Тем не менее, над перегородкой кровь смешанная, так что пресмыкающиеся, как и земноводные,
относятся к пойкилотермным (холоднокровным) животным.
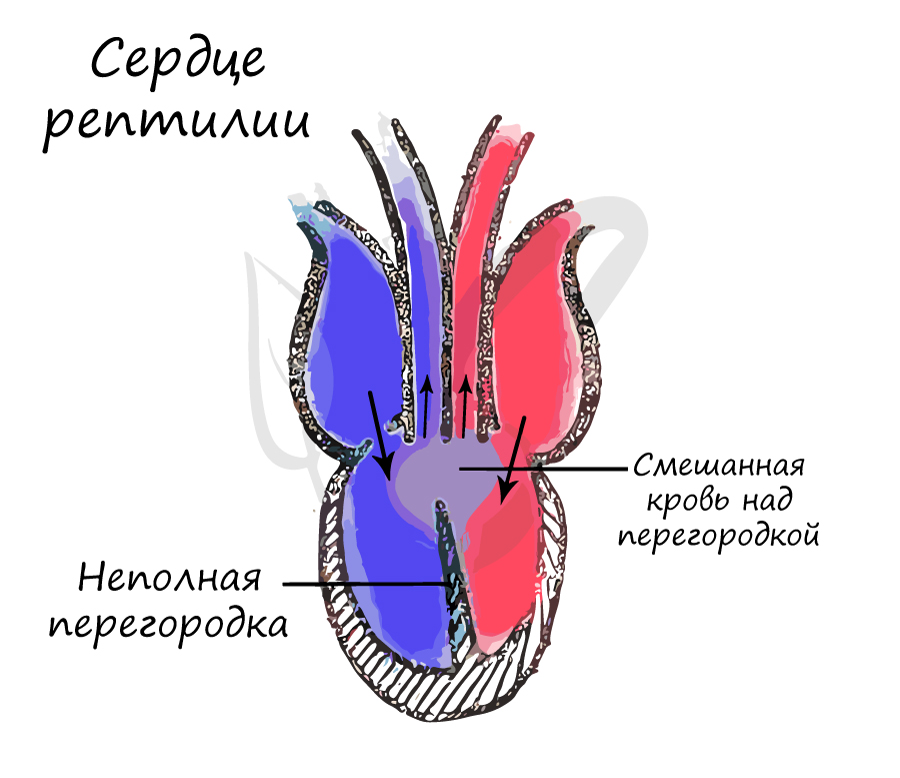
У пресмыкающихся впервые возникают зачатки новой коры головного мозга, совершенствуется воспринимающая и интегрирующая
функции головного мозга. Становится возможным более сложное поведение.
Возникает тазовая (вторичная) почка – метанефрос, выделительный каналец которой гораздо длиннее: становится возможным обратное
всасывание веществ. Засушливый климат, в котором живут рептилии, располагает к экономии воды, их моча становится
более концентрированной.
Отряды пресмыкающихся

В составе класса можно выделить 4 отряда, каждый из которых мы вкратце обсудим. Рептилии традиционно изучаются на примере
типичного представителя – прыткой ящерицы, входящей в состав отряда чешуйчатые. С него мы и начнем знакомство с рептилиями.
Отряд чешуйчатые – прыткая ящерица
- Покровы, опорно-двигательная система
- Пищеварительная система
- Дыхательная система
- Кровеносная система
- Выделительная система
- Нервная система
- Половая система
Тело ящерицы покрыто сухой кожей, практически лишенной желез, с роговыми чешуями и щитками. Такое строение
покровов тела предотвращает высыхание организма, защищает от потери воды. Испарение через кожу
сохраняется, но в минимальном объеме.

Тело подразделяется на голову, туловище, хвост и две пары конечностей. Конечности расположены по бокам туловища, поэтому
поднять голову высоко над землей пресмыкающиеся не могут. Конечности пятипалого типа, перепонки между пальцами отсутствуют.
Предполагаю, вам с детства известно о том, что ящерицы могут отбрасывать свои хвосты. Это действительно так, для
ящериц характерна автотомия (от греч. autos – сам и греч. tome – отсечение) – способность животного самопроизвольно
отбрасывать части своего тела.
В случае нападения
хищника автотомия может спасти жизнь ящерицы, так как отброшенный хвост приковывает к себе внимание и хищник
перестает преследовать ящерицу.

Скелет почти полностью окостеневший, более прочный, чем у земноводных. Позвоночник состоит из 5 отделов: шейный (8 позвонков),
грудной, поясничный, крестцовый и хвостовой. Ребра, сочлененные одним концом с грудными позвонками, а другим – с
грудиной, образуют замкнутую костную структуру – грудную клетку.
К ребрам крепятся впервые возникшие у пресмыкающихся межреберные мышцы, участвующие в дыхании.
За счет наличия 8 шейных позвонков значительно увеличивается подвижность головы, что имеет крайне важное значения для
добывания пищи и ориентировке в окружающем мире. Полость тела рептилий – целом.

Скелеты поясов конечностей служат опорой для самих конечностей. Передний (плечевой) пояс состоит из парных ключиц, лопаток, вороньих
костей (коракоидов) и надгрудинника (надключицы), а также хрящей. Задний (тазовый) пояс конечностей образован тремя костями: седалищной,
лобковой и подвздошной, также включает в себя хрящевые прослойки между ними.
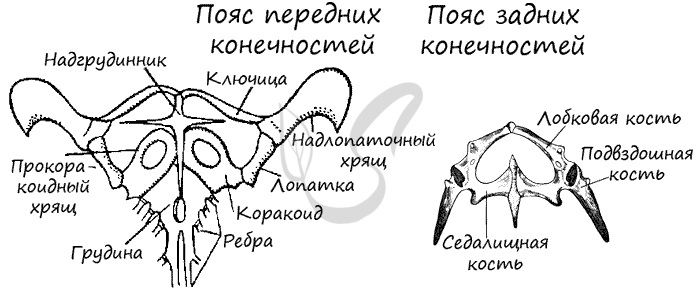
Скелеты передних и задних свободных конечностей аналогичны по строению конечностям амфибий. Однако стоит заметить, что
голень и предплечье не состоят из одной единой кости: они подразделяются на лучевую и локтевую (составляют предплечье),
малоберцовую и большеберцовую (составляют голень).

Строение пищеварительной системы рептилий более дифференцировано по сравнению с таковым у земноводных.
Конические зубы прочно срастаются с челюстями.
Хорошо развиты слюнные железы, которые участвуют не только в смачивании пищи (как у амфибий), но и ферментативно обрабатывают пищевой комок за счет фермента – амилазы, расщепляющей углеводы.
Ротовая полость переходит в короткую глотку, которая продолжается в пищевод. Пищевод расширяется и переходит в желудок,
от которого начинается тонкий, а затем толстый отдел кишечника. В тонкую кишку открываются протоки печени, поджелудочной
железы и желчного пузыря.
Граница между тонкой и толстой кишкой особенно хорошо выражена: на ее месте располагается зачаток
слепой кишки. Оканчивается пищеварительная система клоакой.
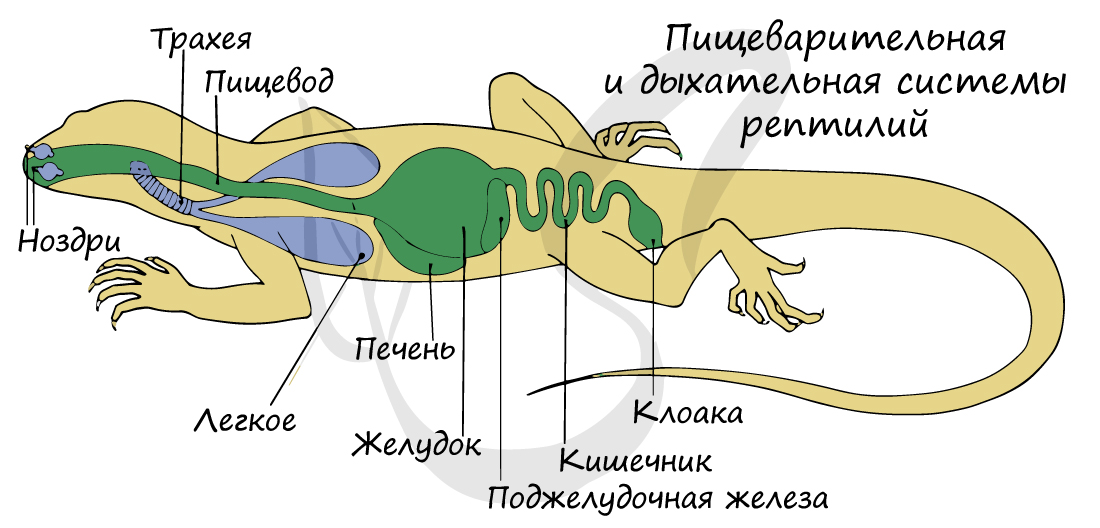
Кожа не участвует в дыхании, эту функцию берут на себя возникшие у рептилий ячеистые легкие. По сравнению с легочными
мешками амфибий, ячеистые легкие имеют бо́льшую дыхательную поверхность.
Воздух присасывается в легкие благодаря движениям грудной клетки, которые происходят за счет сокращения межреберных
мышц. Через воздухоносные пути: хоаны, глотку, гортань, трахею и бронхи – воздух достигает пузырьков, альвеол,
стенка которых густо оплетена капиллярами, в которых и происходит газообмен.
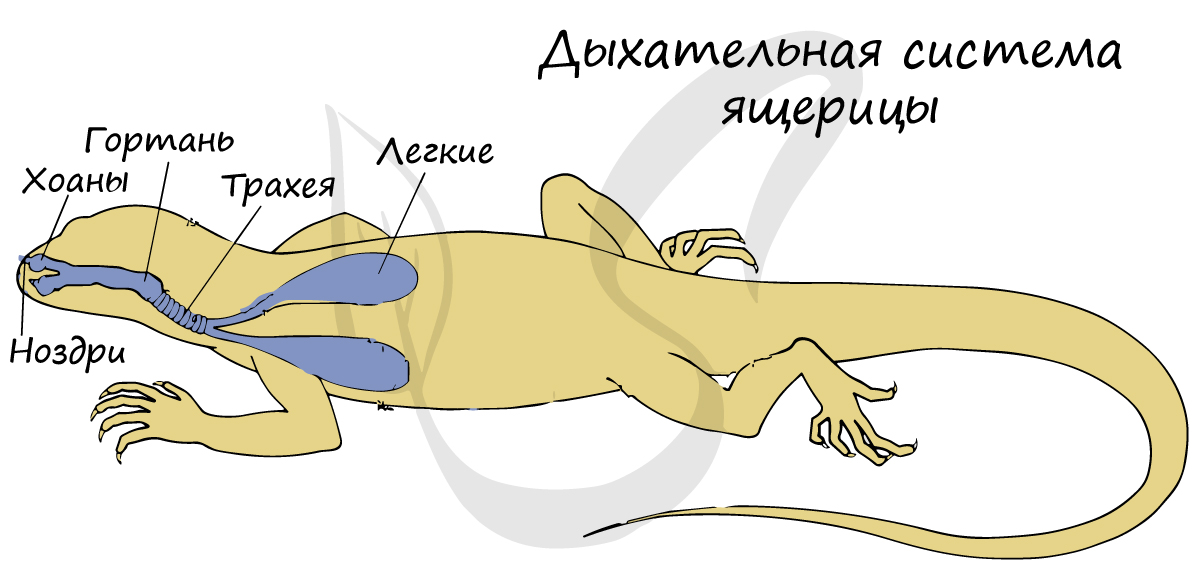
Поскольку рептилии холоднокровные животные, то интенсивность их обмена веществ напрямую зависит от температуры окружающей
среды. Частота дыхательных движений также коррелирует с метаболизмом, поэтому чем выше температура окружающей среды, тем
чаще дышит животное.
Кровеносная система замкнутая. Сердце трехкамерное: два предсердия и один желудочек с неполной перегородкой.
Разделение крови в желудочке эффективнее,
чем у земноводных, но над перегородкой кровь смешанная, поэтому рептилии – холоднокровные.
От сердца отходят 3 сосуда: легочная артерия, по которой венозная кровь поступает в легкие, где насыщается кислородом,
и две дуги аорты – правая и левая. От правой дуги аорты отходят сонные артерии, направляющиеся к головному мозгу.
Правая дуга аорты начинается от левой части желудка
– вследствие чего у рептилий анатомически обусловлено поступление более насыщенной кислородом – артериальной крови – к
головному мозгу, остальные органы получают смешанную кровь из левой дуги аорты.
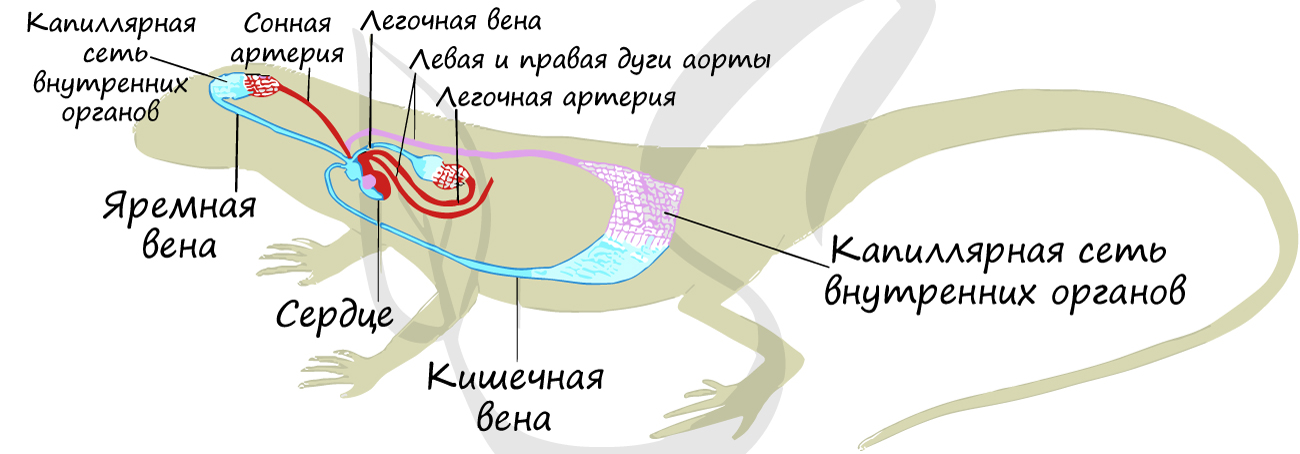
По сравнению с земноводными выделительная система у рептилий более прогрессивного строения, представлена тазовыми (вторичными) почками, называемыми также – метанефрос, позволяющими гораздо эффективнее удерживать воду в организме, препятствуя ее избыточному удалению.
Выделительные канальцы становятся длиннее, за счет чего реабсорбция происходит лучше: из их просвета большее количество воды успевает обратно всосаться
в кровеносное русло. Это нужно для того, чтобы удержать воду в организме, ведь рептилии обитают в засушливых местах. Конечный продукт
обмена веществ у пресмыкающихся – мочевая кислота.
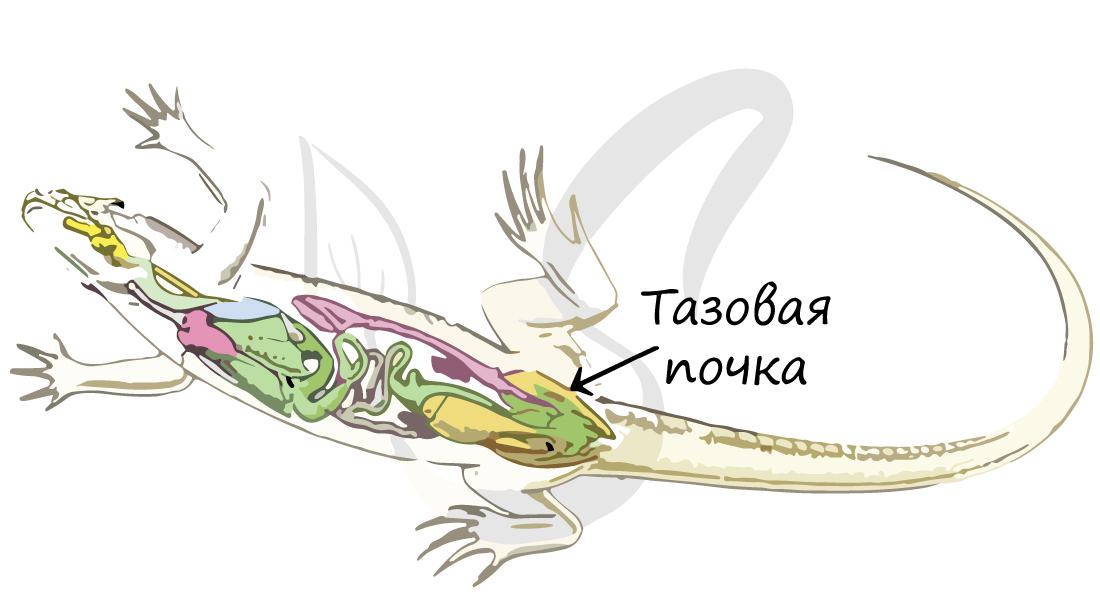
Сравнивая головной мозг земноводных и пресмыкающихся, становятся заметны несколько существенных отличий.
Мозжечок развит
лучше, чем у земноводных. У рептилий появляются зачатки новой коры переднего мозга, он значительно увеличивается в объеме
и перестает быть исключительно обонятельным центром, вбирая в себя новые функции.
Для рептилий характерно более сложное (чем для амфибий) поведение, помимо
заложенных генетически безусловных рефлексов, они способны вырабатывать адаптивные, условные рефлексы.
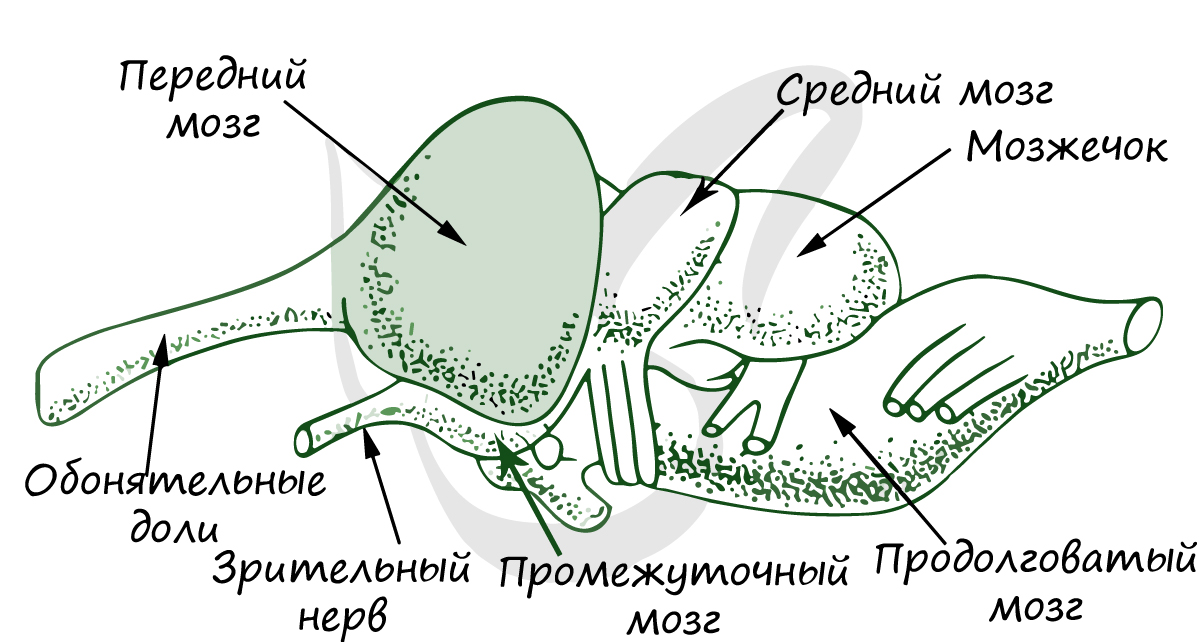
Органы чувств также претерпевают изменения. Аккомодация, настройка глаза на наилучшее видение объекта, теперь достигается не только
перемещением хрусталика вперед-назад (так было у амфибий), но и изменением кривизны хрусталика. Это называется – двойная аккомодация,
настройка глаза на резкость, так что видят рептилии лучше амфибий.
Веки и мигательная перепонка служат для увлажнения поверхности глаза и предохраняют от высыхания.

Орган слуха также совершенствуется: появляется круглое окно во внутреннем ухе, улучшается передача звуковых колебаний, благодаря
чему слух рептилий становится острее. Среднее ухо по-прежнему представлено стремечком и барабанной перепонкой.

Имеются слаборазвитые органы вкуса, локализующиеся в глотке, и органы тепловой чувствительности, особенно хорошо развитые у змей,
расположенные между глазом и носом на лице. Органы обоняния находятся в хоанах. Хоаны – внутренние носовые отверстия, с помощью
которых полость носа сообщается с глоткой.
Мужская половая система представлена семенниками и отходящими от них семяпроводами, впадающими в мочеточники. Мочеточники открываются в клоаку. У самцов имеются особые совокупительные органы, предназначенные для введения семенной жидкости
в половые пути самки. Оплодотворение у рептилий внутреннее.
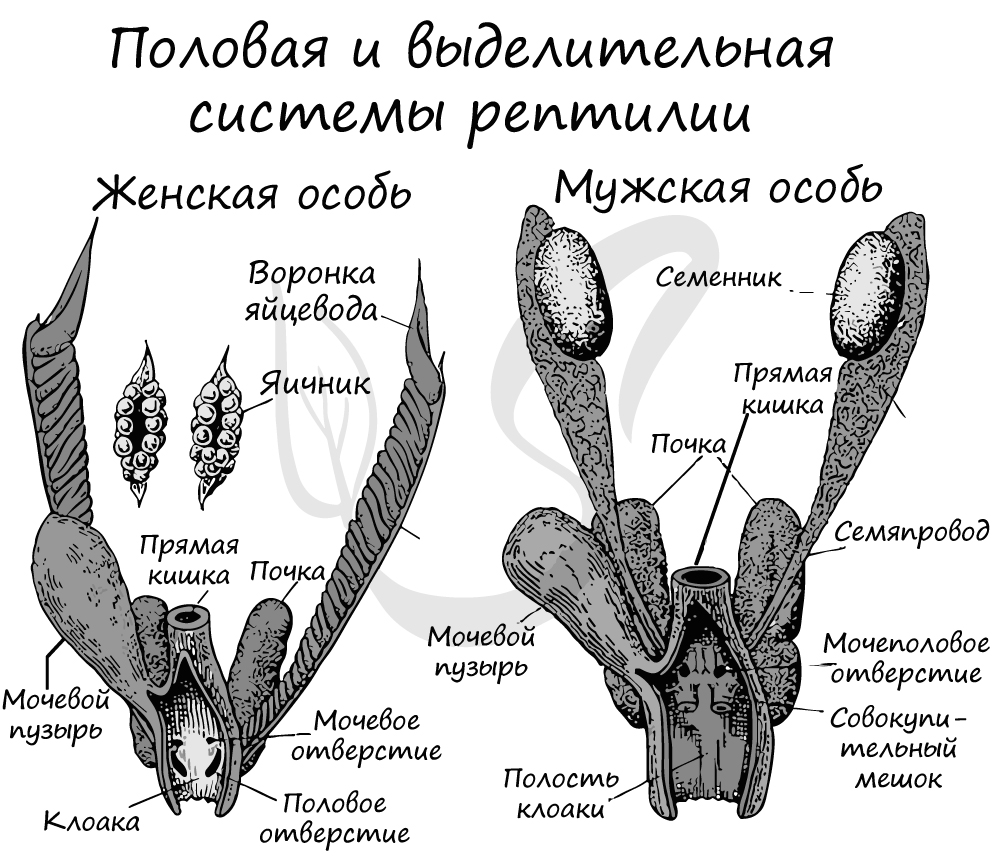
У самок половая система состоит из яичников, от которых отходят яйцеводы, впадающие в клоаку. Сперматозоиды оплодотворяют
яйцеклетки в верхней части яйцевода.
По мере продвижения яйца вниз по яйцеводу оно покрывается сначала белковой оболочкой,
а затем скорлуповой. Эти оболочки – приспособление к жизни на суше, обеспечивающие всестороннюю защиту зародыша.
Напомню, что пресмыкающиеся (также, как птицы и млекопитающие) относятся к группе амниот, зародыш у которых окружен зародышевыми оболочками:
наружной – серозой, внутренней – амниотической. Имеется особый зародышевый орган дыхания – аллантоис.
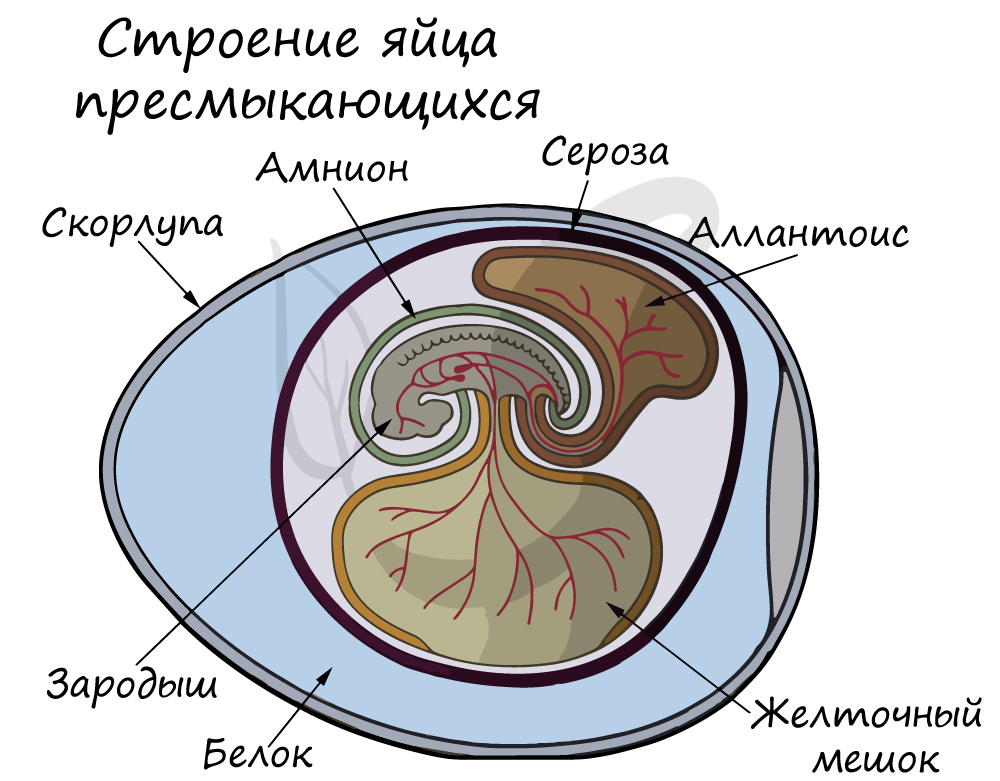
На этом наша беседа о прыткой ящерице подходит к концу. Впереди ждут другие представители рептилий. Из отряда чешуйчатых, помимо
прыткой ящерицы, хочется отметить гекконов – мелких примитивных ящериц, ведущих ночной образ жизни, отлично лазающих по деревьям,
скалам и стенам домов. Самые крупные представители чешуйчатых – вараны, обитающие в Южной Азии, Африке, – достигают в длину до 3,5 метров.

Змеи также принадлежат к отряду чешуйчатые. Это безногие животные, адаптировавшиеся к перемещениям в густом лесном покрове,
кронах деревьев. У змей взгляд немигающий: отсутствуют подвижные веки. Однако их отсутствие не означает, что поверхность глаз не
нуждается в увлажнении: у них имеется мигательная перепонка, смачивающая поверхность глаза.

Змеи подразделяются на ядовитых и неядовитых. Ядовитыми являются гадюка обыкновенная, песчаная эфа, черная мамба, кобра,
гюрза, гремучие и морские змеи. К неядовитым относятся полозы, ужи, удавы.

Черепахи
Имеют уплощенное тело, покрытое панцирем из двух щитков: брюшного и спинного. Примечательно, что на задних конечностях
имеются межпальцевые перепонки для плавания. Морские черепахи продвинулись еще дальше: их задние и передние конечности видоизменены
в ласты, которые служат приспособлением к водному образу жизни.
Есть растительноядные и хищные формы. Настоящих зубов у черепах нет, их челюсти снабжены режущим роговым краем – клювом.

Крокодилы
Крокодилы – еще один отряд класса рептилий. Крокодилы населяют реки и озера тропических стран. Это крупные животные,
тело которых покрывают роговые чешуи. На задних конечностях крокодилы имеют межпальцевые перепонки. Зубы остроконечные, крупные,
располагаются в альвеолах челюсти, как и у млекопитающих.
Легкие имеют более прогрессивное строение – губчатое. Кровеносная система включает в себя четырех (!) камерное сердце,
кровь в сердце не смешивается, так как желудочек полностью разделяется межжелудочковой перегородкой на две камеры.
Однако, крокодилы все равно остаются холоднокровными животными: кровь смешивается в спинной аорте, образованной в
результате слияния левой и правой дуги аорты.

Вероятно, с детства многим известна история про птицу – крокодилова сторожа, которая питается остатками пищи из пасти крокодила.
Спешу сообщить, что это скорее миф, чем правда. Крокодилы вовсе не нуждаются в подобной “чистке”, а птицы предпочитают
питаться насекомыми, в том числе мухами, которые слетаются на остатки мяса в пасти крокодила.
Клювоголовые
Мы добрались до самого древнего отряда рептилий. На сегодняшний день остался единственный его представитель – гаттерия,
обитающая на нескольких островах Новой Зеландии. По внешнему виду напоминает игуану, ведет ночной образ жизни, питается насекомыми,
моллюсками, червями. Гаттерия является вымирающим видом.

Значение пресмыкающихся
Как и все живые организмы, пресмыкающиеся являются звеном в цепи питания (консументами). Регулируют численность насекомых,
питаясь ими. Среди рептилий встречаются как хищники, так и растительноядные формы.
Рептилии имеют пищевое значение. В Африке и Азии употребляют в пищу около сотни различных блюд из мяса змей;
яйца, жир и мясо морской черепахи также употребляются в пищу. Из кожи крокодилов изготавливают различные изделия.

© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Пресмыкающиеся — первые настоящие наземные позвоночные. Земноводных нельзя считать истинно наземными, потому что они не могут размножаться без воды, в то время как пресмыкающиеся способны откладывать яйца даже в пустыне. Некоторые рептилии вторично вернулись в водную среду обитания.
Класс насчитывает около 8 000 видов. Самые крупные наземные животные, динозавры, были пресмыкающимися.
Следующие изменения в строении тела позволили рептилиям выйти на сушу:
- Сухая, ороговевшая кожа. Она не пропускает через себя газы и воду, препятствуя испарению;
- Лёгкие и дыхательные пути, которые устроены сложнее, чем у земноводных;
- Частичное разделение кровотока сердца (с помощью перегородки), что делает кровоснабжение более эффективным;
- Внутренне оплодотворение и защитные влагонепроницаемые оболочки яйца позволяют зародышу развиваться на суше.
Тело и покровы
Верхний слой кожи состоит из ороговевших чешуй, которые регулярно слущиваются. Нижний слой – живой, он обеспечивает образование новых чешуй. Ороговевший эпителий защищает от механических повреждений и иссушения.
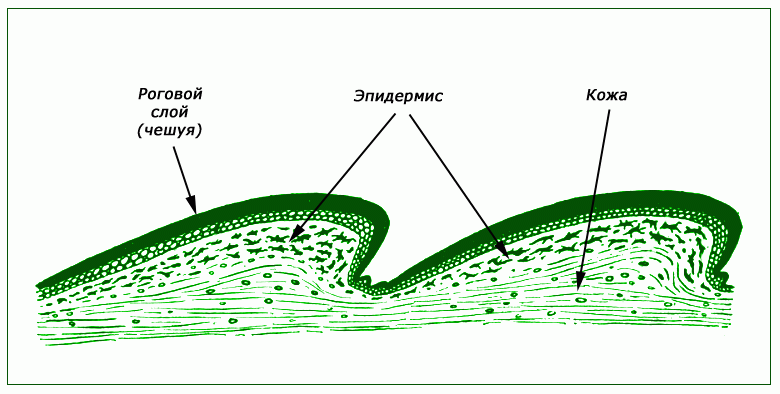
Тело состоит из головы, туловища и хвоста (у некоторых видов хвост отсутствует). Задняя конечность делится на бедро, голень и стопу, передняя – на плечо, предплечье и кисть.
Опорно-двигательная система
Шейный отдел позвоночника развит хорошо, состоит из 8 позвонков, отчего шея становится подвижной по сравнению с земноводными.
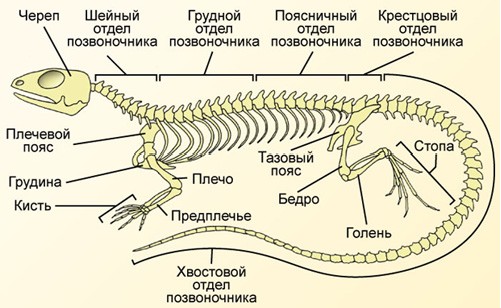
От позвонков грудного отдела отходят рёбра. У рептилий рёбра срастаются спереди, появляется настоящая грудная клетка. С её помощью животное может регулировать объём лёгких, активно дышать. Грудная клетка черепах образует каркас для массивного панциря. У змей может быть до 150 пар рёбер, при этом они не срастаются. Это позволяет змеям глотать крупную пищу.
Конечности расположены по бокам туловища, а не под ним, за что пресмыкающиеся и получили своё название. Во втором отделе конечностей, в отличии от скелета земноводных, по две кости: лучевая и локтевая в передних, большая и малая берцовые в задних конечностях.
Мышечная система более разнообразна, если сравнивать с земноводными. Появляется межрёберная мускулатура, она участвует в акте дыхания.
Пищеварительная система
Типичная, состоит из желудочно-кишечного тракта, печени, поджелудочной железы. Наружу кишечник открывается клоакой, половая, пищеварительная и выделительная системы имеют совместный выход.
Кровеносная система
Имеется два круга кровообращения: большой (через тело) и малый (к лёгким). От сердца отходят две дуги аорты. Сердце трёхкамерное, но между желудочками есть неполная перегородка. Исключение составляют крокодилы, имеющие четырёхкамерное сердце, у них круги кровообращения разделены полностью.
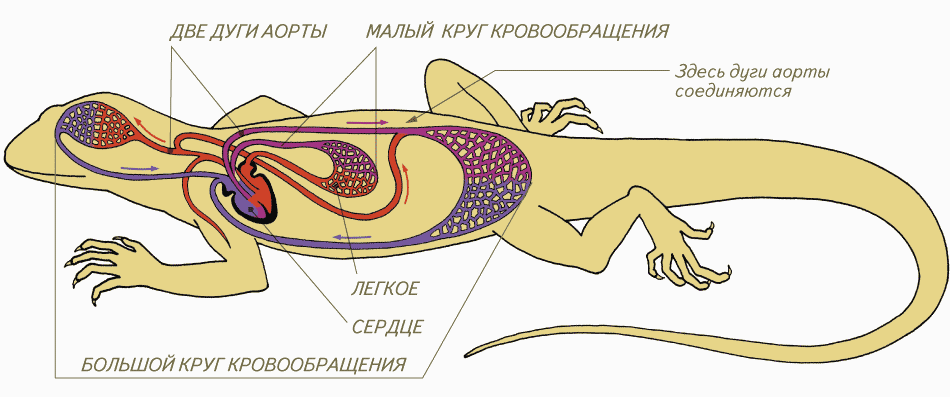
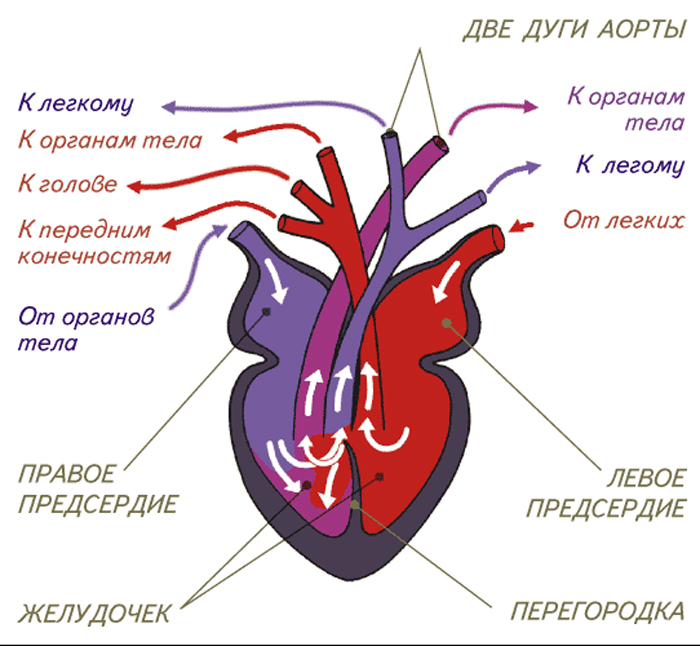
От лёгких артериальная кровь течёт в левое предсердие, от него попадает в левый желудочек. Между левым и правым желудочками неполная перегородка, поэтому часть артериальной крови смешивается с венозной. При этом к голове и передним конечностям идёт артериальная кровь, а к телу, внутренним органам, задним конечностям – смешанная.
Кровеносная система замкнутая, все форменные элементы крови имеют ядра.
Дыхательная система
Лёгкие плотные, плохо растяжимы. Площадь поверхности лёгких увеличена за счёт губчатого строения, наличия множества перегородок. Нагнетание воздуха происходит при движении рёбер.
Змеи обладают одним длинным лёгким, расположенным вдоль тела.
Выделительная система
Представлена почками, причём почки не туловищные, как у амфибий, а смещаются к тазу. Тазовые почки в силу своего строения лучше сохраняют воду. Воронка отсутствует, поэтому не происходит фильтрации тканевой жидкости, все вещества поступают в почки исключительно из крови.
Система извитых канальцев позволяет возвратить из первичной мочи полезные вещества и воду. Концентрированная мочевая кислота по мочеточникам выводится в клоаку. Пустынные пресмыкающиеся могут обходиться без питья, получая необходимую жидкость из добычи.
Нервная система
Хорошо развит мозжечок, что позволяет совершать разнообразные сложные движения. Достаточно развиты полушария переднего мозга, это подтверждается сложным поведением пресмыкающихся. Появляется кора больших полушарий. Она тонкая, гладкая, не имеет извилин.
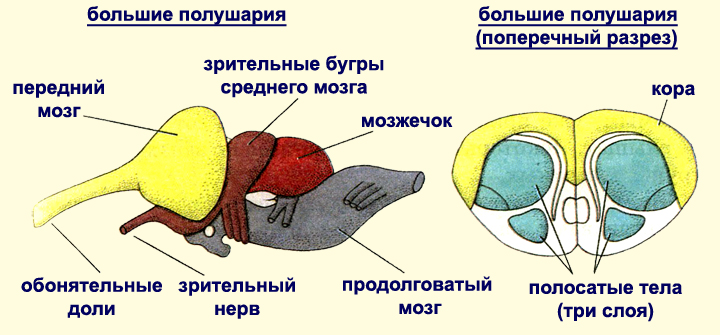
Ведущее чувство рептилий – зрение. Глаза хорошо приспособлены к работе в воздушной среде. Слёзные железы предохраняют глаз от высыхания, защитную функцию выполняют наружные веки и мигательная перепонка. Зрение может быть цветным или чёрно-белым. Присутствует также теменной глаз – группа светочувствительных рецепторов под кожей. Он анализирует освещённость, участвует в синхронизации суточных ритмов.
Орган слуха близок по устройству к уху земноводных. На поверхности кожи находится барабанная перепонка, за ней – среднее ухо со слуховой косточкой (стремечко) и внутреннее ухо.
Органы обоняния развиты лучше, чем у земноводных. Ящерицы могут по запаху находить добычу под песком. В глотке рептилий расположены вкусовые луковицы. Осязание развито хорошо, в качестве органов осязание часто используется язык.
Уникальный орган чувств – термолокатор, он воспринимает инфракрасное (тепловое) излучение, хорошо развит у некоторых змей.
Размножение
Рептилии раздельнополы. Половые органы самца – парные семенники, лежащие в полости тела по бокам от позвоночника. Сперма попадает наружу через клоаку, у многих видов есть копулятивный орган. У самок имеются парные яичники, скорлуповые железы. Оплодотворение внутреннее, развитие без превращения.
После спаривания яйцеклетка растёт и обволакивается несколькими белковыми оболочками. Наружная оболочка – твёрдая скорлупная. Самка может ухаживать за кладкой яиц, некоторые крокодилы заботятся о вылупившихся детёнышах.
Многообразие, роль в природе
Современные представители класса пресмыкающихся – несколько разрозненных групп, оставшихся после Мезозоя, времени расцвета рептилий. Класс делят на четыре отряда: чешуйчатые (6500 видов), черепахи (230 видов), крокодилы (около 20 видов) и клювоголовые (1 вид – гаттерия).
Чешуйчатые – процветающий отряд, включающий ящериц, змей и хамелеонов. Змеи отличаются отсутствием конечностей, плечевого пояса, подвижным сочленением челюстей. У некоторых змей есть ядовитые железы. Змеи способны линять: одномоментно освобождаться от старого слоя ороговевшего эпителия, а заодно и от паразитов.
Черепахи имеют крупный панцирь, в который могут спрятать голову и конечности. Структура панциря способна выдержать вес, превосходящий в 200 раз вес самого животного. Зубов нет, первичная обработка пищи происходит с помощью клюва. Условно отряд можно разделить на морских и наземных черепах. Морские черепахи известны своим долгожительством, они могут жить до 150 лет.
Крокодилы являются полуводными животными, они населяют прибрежные и материковые тропические водоёмы. Крокодилы обладают мощными вытянутыми челюстями и множеством сменяющихся в течении жизни зубов. Глаза и ноздри могут выступать над поверхностью воды, в то время как остальное тело остаётся незаметным. Это позволяет крокодилам подкарауливать добычу.
Как и все животные, рептилии участвуют в пищевых цепях. Они могут питаться вредными насекомыми, грызунами.
Змеиный яд используют в медицинской практике. Мясо змей, ящериц и черепах люди употребляют в пищу. Из панцирей черепах делают украшения, предметы быта. Кожа крокодилов и змей используется для изготовления одежды, обуви, аксессуаров. Это ведёт к вымиранию некоторых видов.
Задание EB11103
Какие черты в строении рептилий указывают на приспособление к жизни на суше?
- Сухая кожа без желез препятствует потере воды.
- Внутреннее оплодотворение. Яйца покрытые скорлупой с большим количеством питательных веществ.
- Легочное дыхание (мелкоячеистые легкие).
или, Увеличивается дыхательная поверхность легких. Неполная перегородка в желудочке уменьшает смешивание крови. – Всё это позволяет доносить до органов кровь, богатую кислородом.Ответ: см. решение
pазбирался: Ксения Алексеевна | обсудить разбор
Задание EB10412
Установите соответствие между видом животного и особенностью строения его сердца.
| ВИД ЖИВОТНОГО | СТРОЕНИЕ СЕРДЦА |
|
А) прыткая ящерица Б) обыкновенный тритон В) озёрная лягушка Г) синий кит Д) серая крыса Е) сокол сапсан |
1) трёхкамерное без перегородки в желудочке 2) трёхкамерное с неполной перегородкой в желудочке 3) четырёхкамерное |
Четырехкамерное сердце у птиц и млекопитающих. Трехкамерное без перегородки — у земноводных. Трехкамерное с перегородкой — у рептилий.
PS: киты — млекопитающие, тритоны — земноводные.
Ответ: 211333
pазбирался: Ксения Алексеевна | обсудить разбор
Задание EB10408
Для каждого животного выберите температуру тела.
| ЖИВОТНЫЕ | ТЕМПЕРАТУРА |
|
А) Речной окунь Б) Голубая акула В) Заяц–беляк Г) Серая жаба Д) Большая синица Е) Гренландский тюлень Ж) Прыткая ящерица |
1) постоянная 2) непостоянная |
Непостоянная температура тела наблюдается у беспозвоночных и у некоторых из позвоночных: рыбы, земноводные и рептилии.
Ответ: 2212112
pазбирался: Ксения Алексеевна | обсудить разбор
Задание EB10406
Соотнесите признаки животных с классами, для которых этот признак характерен.
| ПРИЗНАКИ ЖИВОТНЫХ | КЛАССЫ |
|
А) оплодотворение внутреннее Б) оплодотворение у большинства видов наружное В) непрямое развитие Г) размножение и развитие происходит на суше Д) тонкая кожа, покрытая слизью Е) яйца с большим запасом питательных веществ |
1) земноводные 2) пресмыкающиеся |
Разберемся с названиями представленных классов:
Земноводные, два корня. Живут представители данного класса и на суше, и в воде. Ярчайший пример — лягушка. Она сидит и в воде, и на кувшинках.
Пресмыкающиеся. Название подобрано не просто так. Эти животные «пресмыкаются» к земле. Когда они передвигаются, то часть тела волочится по земле. По-другому этот класс именуется «рептилии».
Вот мы и прикинули, какой класс что из себя представляет. Выберем по представителю. Пусть при размышлениях о земноводных мы будем думать о лягушках, а при мыслях о пресмыкающихся – ящерица или крокодил.
Начнем с простого: вариант с кожей.
У ящерицы кожа плотная, иногда как будто с жесткими чешуями, а у лягушки кожа тоненькая, склизкая. Один ответ уже есть: Д — 1.
Теперь подумаем о том, какие как развивается потомство данных животных.
У лягушек что-то похожее на икру, а вот у ящериц яйца скорее походи на куриные, тоже с твердой скорлупой, крупные, с желтком. В яйце развивается плод, ему нужно питание, которое он получает из желтка.
У кого же будет больше питательных веществ? Наверняка у рептилий. Е — 2.
Подумаем про оплодотворение. У лягушек очень много этой «икры». Кстати, как оплодотворяется икра? В воде. Самки ее мечут, она остается на растениях и прочих поверхностях дна, а самцы потом ее осеменяют. Не вся икра выживет, часть ведь съедят обитатели водоема еще неоплодотворенной, часть куда-нибудь отнесет. В общем, известный факт, который легко понять: чем больше количество потомства, тем меньше забота о нем, тем ниже уровень организации. У лягушек количество отложенных яиц большое, у рептилий-несколько штук.
В воде у лягушек изначально неоплодотворенные яйца. Рептилии же откладывают уже оплодотворенные яйца. Значит, у лягушек оплодотворение наружное, а у пресмыкающихся- внутреннее. А — 2, Б — 1.
Рассмотрим вариант про развитие и размножение в воде. Ну вот у лягушки размножение и развитие, т.е головастики, в воде. А у рептилий? Пожалуйста, не думайте, например, о крокодилах сейчас. Да, часто его можно увидеть в воде, что может натолкнуть на мысли. Но вот подумайте о ящерках на полянках, в лесу. Не бегают же они размножаться в водоем? Вряд ли, да и яйца с большим количеством желтка, скорлупой и зародышем просто потонут. Г — 2.
Теперь все же теория:
Прямое развитие — рождение потомства, внешне похожего на взрослый организм.
Непрямое развитие (личиночное развитие, развитие с метаморфозом) — появившийся организм отличается по строению от взрослого.
У ящерицы появляется маленькая ящерка, у крокодила — крокодильчик, а у лягушки что-то непонятное под названием головастик. Так вот, он отличается от взрослой особи, это и есть непрямое развитие. В — 1.
Ответ: 211212
pазбирался: Ксения Алексеевна | обсудить разбор
Задание EB20519
Какие из перечисленных ароморфозов привели к возникновению рептилий? Выберите три верных ответа из шести и запишите в таблицу цифры, под которыми они указаны.
- появление грудной клетки для засасывания воздуха в легкие
- возникновение покрова из ороговевших чешуй
- формирование пятипалых конечностей
- появление оболочек яиц
- возникновение кожного дыхания
- появление второго круга кровообращения
Ароморфоз — такое изменение в организме, что оно позволило шагнуть на следующую ступень развития. У каждого класса свои ароморфозы. Что самое главной сделали рептилии? Конечно, они вышли на сушу, для чего, в первую очередь, нужно было дышать только воздухом.
Теперь, когда жизнь вышла на сушу, необходима защита, поэтому покровы тела меняются, с мягких, покрытых слизью на сухие и ороговевшие.
Пятипалые конечности были и до рептилий, посмотрите на лягушку.
Для успешного распространения нужно выходить потомство. В условиях суши нужна защита, то есть оболочки для яиц. Иначе, они попросту высохнут, не говоря о том, что легко травмируются или будут съедены.
Кожное дыхание было и у земноводных.
Второй круг кровообращения так же есть еще у земноводных.
Ответ: 124
pазбирался: Ксения Алексеевна | обсудить разбор
Задание EB22081
Назовите отличительные признаки пресмыкающихся от птиц? Выберите три верных ответа из шести и запишите цифры, под которыми они указаны.
- смешанность крови
- непостоянная температура тела
- сухой роговой покров
- медленный обмен веществ
- внутреннее оплодотворение
- два круга кровообращения
У птиц сердце четырехкамерное, кровь уже делится на артериальную и венозную, в отличии от пресмыкающихся, у которых еще трехкамерное сердце, а кровь еще смешивается.
У птиц и млекопитающих температура тела постоянная.
И у рептилий, и у птиц покровы очень сухие.
Обмен веществ. Вспомните колибри, она ведь ест с десяток раз на дню. А питон? Заглотил свою жертву и лежит несколько дней, переваривает.
Оплодотворение внутренние у обоих классов.
Хоть кровь и смешивается, а круга два.
Ответ: 134
pазбирался: Ксения Алексеевна | обсудить разбор
Даниил Романович | Просмотров: 17.3k
